Dominacja (genetyka)

W genetyce dominacja jest zjawiskiem polegającym na tym, że jeden wariant ( allel) genu na chromosomie maskuje lub zastępuje wpływ innego wariantu tego samego genu na drugą kopię chromosomu . Pierwszy wariant jest określany jako dominujący , a drugi recesywny . Ten stan posiadania dwóch różnych wariantów tego samego genu na każdym chromosomie jest pierwotnie spowodowany mutacją w jednym z genów, nową ( de novo ) lub odziedziczoną . Terminy autosomalny dominujący lub autosomalny recesywny są używane do opisania wariantów genów na chromosomach niezwiązanych z płcią ( autosomy ) i związanych z nimi cech, podczas gdy te na chromosomach płciowych ( allosomy ) są określane jako dominujący sprzężony z chromosomem X , recesywny sprzężony z chromosomem X lub sprzężony z Y ; mają one wzór dziedziczenia i prezentacji, który zależy od płci zarówno rodzica, jak i dziecka (patrz Powiązanie płci ). Ponieważ istnieje tylko jedna kopia chromosomu Y , cechy sprzężone z Y nie mogą być dominujące ani recesywne. Ponadto istnieją inne formy dominacji, takie jak dominacja niepełna , w której wariant genu ma częściowy wpływ w porównaniu z sytuacją, gdy jest obecny na obu chromosomach, oraz współdominacja , w której różne warianty na każdym chromosomie wykazują powiązane cechy.
Dominacja jest kluczowym pojęciem w dziedziczeniu mendlowskim i genetyce klasycznej . Litery i kwadraty Punnetta są używane do zademonstrowania zasad dominacji w nauczaniu, a używanie wielkich liter dla alleli dominujących i małych dla alleli recesywnych jest powszechnie stosowaną konwencją. Klasycznym przykładem dominacji jest dziedziczenie kształtu nasion grochu . Groch może być okrągły, związany z allelem R lub pomarszczony, związany z allelem r . W tym przypadku możliwe są trzy kombinacje alleli (genotypów): RR , Rr i rr . Osobniki RR ( homozygotyczne ) mają okrągły groszek, a osobniki rr ( homozygotyczne ) mają pomarszczony groszek. U osobników Rr ( heterozygotycznych ) allel R maskuje obecność allelu r , więc te osobniki również mają okrągły groszek. Zatem allel R dominuje nad allelem r , a allel r jest recesywny w stosunku do allelu R.
Dominacja nie jest związana z allelem ani jego cechami ( fenotypem ). Jest to efekt ściśle względny między dwoma allelami danego genu o dowolnej funkcji; jeden allel może być dominujący nad drugim allelem tego samego genu, recesywny w stosunku do trzeciego i współdominujący z czwartym. Dodatkowo jeden allel może dominować dla jednej cechy, ale nie dla innych.
Dominacja różni się od epistazy , zjawiska, w którym allel jednego genu maskuje działanie alleli innego genu .
Tło
_(14582377398).jpg)
Pojęcie dominacji wprowadził Gregor Johann Mendel . Chociaż Mendel, „ojciec genetyki”, po raz pierwszy użył tego terminu w latach sześćdziesiątych XIX wieku, nie był on powszechnie znany aż do początku XX wieku. Mendel zauważył, że dla różnych cech grochu ogrodowego, związanych z wyglądem nasion, strąków nasiennych i roślin, istniały dwa oddzielne fenotypy, takie jak okrągłe i pomarszczone nasiona, żółte i zielone nasiona, czerwone i białe kwiaty lub rośliny wysokie i niskie. Hodowane oddzielnie rośliny zawsze wytwarzały te same fenotypy, pokolenie po pokoleniu. Jednakże, kiedy linie o różnych fenotypach zostały skrzyżowane (krzyżowane), jeden i tylko jeden fenotyp rodzicielski pojawił się u potomstwa (zielony, okrągły, czerwony lub wysoki). Jednakże, kiedy te hybrydowe zostały skrzyżowane, rośliny potomne wykazywały dwa oryginalne fenotypy, w charakterystycznym stosunku 3:1, przy czym bardziej powszechnym fenotypem był fenotyp macierzystych roślin hybrydowych. Mendel doszedł do wniosku, że każdy rodzic w pierwszej krzyżówce był homozygotą pod względem różnych alleli (jeden rodzic AA, a drugi rodzic aa), że każdy dał jeden allel potomstwu, w wyniku czego wszystkie te hybrydy były heterozygotami (Aa) i że jeden z dwóch alleli w krzyżówce hybrydowej zdominował ekspresję drugiego: A zamaskowany a. Ostateczne skrzyżowanie dwóch heterozygot (Aa X Aa) dałoby potomstwo AA, Aa i aa w stosunku genotypów 1:2:1, przy czym pierwsze dwie klasy wykazują fenotyp (A), a ostatnia wykazuje fenotyp (a) , tworząc w ten sposób stosunek fenotypów 3: 1.
Mendel nie używał terminów gen, allel, fenotyp, genotyp, homozygota i heterozygota, z których wszystkie zostały wprowadzone później. Wprowadził notację wielkich i małych liter odpowiednio dla alleli dominujących i recesywnych, która jest nadal w użyciu.
W 1928 roku brytyjski genetyk populacyjny Ronald Fisher zaproponował, że dominacja działa w oparciu o dobór naturalny poprzez wkład genów modyfikujących . W 1929 roku amerykański genetyk Sewall Wright odpowiedział, stwierdzając, że dominacja jest po prostu fizjologiczną konsekwencją szlaków metabolicznych i względnej konieczności zaangażowanego genu. Wyjaśnienie Wrighta stało się faktem w genetyce, a debata została w dużej mierze zakończona. Jednak niektóre cechy mogą mieć wpływ na dominację mechanizmów ewolucyjnych.
Chromosomy, geny i allele
Większość zwierząt i niektóre rośliny mają sparowane chromosomy i są opisywane jako diploidalne . Mają dwie wersje każdego chromosomu, jedną wniesioną przez komórkę jajową matki , a drugą przez nasienie ojca , znane jako gamety , opisane jako haploidalne i powstałe w wyniku mejozy . Te gamety następnie łączą się podczas zapłodnienia podczas rozmnażania płciowego , tworząc nową zygotę jednokomórkową, która dzieli się wielokrotnie, w wyniku czego powstaje nowy organizm z taką samą liczbą par chromosomów w każdej komórce (niebędącej gametą), jak jego rodzice. W genetyce ssaków zaburzenia autosomalne dominujące mają rodowody , które wykazują pionowy wzór dziedziczenia.
Każdy chromosom z pasującej (homologicznej) pary jest strukturalnie podobny do drugiego i ma bardzo podobną sekwencję DNA ( loci , pojedyncze locus). DNA w każdym chromosomie działa jako seria oddzielnych genów , które wpływają na różne cechy. Zatem każdy gen ma również odpowiedni homolog, który może istnieć w różnych wersjach zwanych allelami . Allele w tym samym locus na dwóch homologicznych chromosomach mogą być identyczne lub różne.
Na przykład grupa krwi człowieka jest określana przez gen ABO, który koduje warianty enzymu tworzącego grupę krwi A, B, AB lub O, znajdującego się na długim lub q ramieniu chromosomu dziewiątego (9q34.2). Istnieją trzy różne allele, które mogą być obecne w tym locus, ale tylko dwa mogą być obecne u każdego osobnika, jeden odziedziczony po matce i jeden po ojcu.
Jeśli dwa allele danego genu są identyczne, organizm nazywa się homozygotą i mówi się, że jest homozygotą w odniesieniu do tego genu; jeśli zamiast tego dwa allele są różne, organizm jest heterozygotą i jest heterozygotą. Skład genetyczny organizmu, albo w jednym locus, albo we wszystkich jego genach łącznie, nazywany jest jego genotypem . Genotyp organizmu, bezpośrednio i pośrednio, wpływa na jego cechy molekularne, fizyczne i inne, które pojedynczo lub zbiorowo nazywane są jego fenotypem . W heterozygotycznych loci genów dwa allele oddziałują ze sobą, tworząc fenotyp. [ potrzebne źródło ]
Rodzaje dominacji
Całkowita dominacja
W przypadku całkowitej dominacji wpływ jednego allelu w genotypie heterozygotycznym całkowicie maskuje wpływ drugiego. Allel, który maskuje, jest uważany za dominujący w stosunku do drugiego allelu, a allel zamaskowany jest uważany za recesywny .
Całkowita dominacja w fenotypie heterozygoty jest nie do odróżnienia od fenotypu dominującej homozygoty. Klasycznym przykładem całkowitej dominacji jest dziedziczenie kształtu nasion (kształtu grochu) u grochu. Groch może być okrągły (związany z allelem R ) lub pomarszczony (związany z allelem r ). W tym przypadku możliwe są trzy kombinacje alleli ( genotypów ): RR, rr, Rr. RR i rr są homozygotami, a Rr jest heterozygotą. Osobniki RR mają okrągły groszek, a osobniki rr pomarszczony groszek. U osobników Rr allel R maskuje obecność allelu r , więc te osobniki również mają okrągły groszek. Zatem allel R jest całkowicie dominujący w stosunku do allelu r , a allel r jest recesywny w stosunku do allelu R.
Niepełna dominacja

Niepełna dominacja (zwana także częściową dominacją , półdominacją , pośrednim dziedziczeniem lub czasami błędną współdominacją w genetyce gadów) występuje, gdy fenotyp genotypu heterozygotycznego różni się od fenotypów genotypów homozygotycznych i często jest pośredni. Wynik fenotypowy często pojawia się jako mieszana forma cech w stanie heterozygotycznym. Na przykład lwiej paszczy jest homozygotyczny pod względem czerwieni lub bieli. Kiedy czerwony homozygotyczny kwiat jest sparowany z białym homozygotycznym kwiatem, w wyniku powstaje różowy kwiat lwiej paszczy. Różowy lwia paszcza jest wynikiem niepełnej dominacji. Podobny rodzaj niepełnej dominacji można znaleźć w roślinie o czwartej godzinie , w której różowy kolor powstaje, gdy krzyżuje się prawdziwych rodziców o białych i czerwonych kwiatach. W genetyce ilościowej , gdzie fenotypy są mierzone i traktowane liczbowo, jeśli fenotyp heterozygoty jest dokładnie pomiędzy (liczbowo) fenotypem dwóch homozygot, mówi się, że fenotyp nie wykazuje żadnej dominacji, tj. dominacja istnieje tylko wtedy, gdy miara fenotypu heterozygoty jest bliżej do jednej homozygoty niż do drugiej.
Gdy rośliny pokolenia F 1 są samozapylone, stosunek fenotypowy i genotypowy pokolenia F 2 będzie wynosił 1:2:1 (czerwony: różowy: biały).
Zobacz hipotezę częściowej dominacji .
Współdominacja



Współdominacja występuje, gdy wkład obu alleli jest widoczny w fenotypie i żaden allel nie maskuje drugiego.
IA Na przykład w systemie grup krwi ABO IB chemiczne modyfikacje glikoproteiny ( antygenu H) na powierzchni komórek krwi są kontrolowane przez trzy allele, z których dwa są współdominujące względem siebie ( , ) i dominujący nad recesywnym i w locus ABO . Allele I A i I B wytwarzają różne modyfikacje. Enzym kodowany przez IA dodaje N-acetylogalaktozaminę do związanego z błoną antygenu H. Enzym IB dodaje galaktozę. Allel i nie powoduje modyfikacji. Zatem I A i I B są dominujące dla i ( oba osobniki I A I A i I A i mają grupę krwi A, a osobniki I B I B i I B i mają grupę krwi B), ale I A I Osoby B mają obie modyfikacje na swoich komórkach krwi, a zatem mają grupę krwi AB, więc mówi się, że allele I A i I B są współdominujące.
Inny przykład występuje w locus składnika beta-globiny hemoglobiny , gdzie trzy fenotypy cząsteczkowe Hb A /Hb A , Hb A /Hb S i Hb S /Hb S można rozróżnić za pomocą elektroforezy białek . (Stan chorobowy wywołany przez genotyp heterozygotyczny nazywany jest cechą anemii sierpowatej i jest łagodniejszym stanem, który można odróżnić od anemii sierpowatej , dlatego allele wykazują niepełną dominację w odniesieniu do anemii, patrz wyżej). W przypadku większości loci genów na poziomie molekularnym oba allele ulegają ekspresji kodominującej, ponieważ oba są transkrybowane do RNA .
Współdominacja, w której produkty alleliczne współistnieją w fenotypie, różni się od dominacji niepełnej, w której ilościowa interakcja produktów alleli daje fenotyp pośredni. Na przykład w kodominacji czerwony kwiat homozygotyczny i biały kwiat homozygotyczny dadzą potomstwo z czerwonymi i białymi plamami. Gdy rośliny pokolenia F1 są samozapylone, stosunek fenotypowy i genotypowy pokolenia F2 będzie wynosił 1:2:1 (czerwony: cętkowany: biały). Stosunki te są takie same, jak w przypadku niepełnej dominacji. Ponownie, ta klasyczna terminologia jest niewłaściwa – w rzeczywistości w takich przypadkach nie należy mówić o dominacji.
Odnoszenie się do powszechnych błędnych przekonań
Dominacja odnosi się do związku między dwiema wersjami genu. Dominująca cecha jest zwykle zgodna z wzorcami dziedziczenia, które można zobaczyć w kwadratach Punnetta. Jeśli dana osoba ma dwie wersje genu, to gen często obserwowany w kolejnych pokoleniach jest uważany za „dominujący”.
W genetyce istnieje kilka nieporozumień, które są dość powszechne. Uważa się, że cecha dominująca jest „silniejsza” i „obezwładnia” cechę recesywną. Zakłada się również, że cechy dominujące są bardziej dziedziczne i bardziej rozpowszechnione w populacji. Pomysł, że dominujące cechy są męskie lub męskie, jest kolejnym powszechnym nieporozumieniem. Pojawienie się tych różnych idei wynika z różnych koncepcji dominacji w warunkach niegenetycznych; takie jak bycie silnym, potężnym i kontrolującym; co różni się od genetycznej koncepcji dominacji.
Dominacja nie określa, czy allel jest szkodliwy, neutralny czy korzystny. Jednak selekcja musi działać na geny pośrednio poprzez fenotypy, a dominacja wpływa na ekspozycję alleli w fenotypach, stąd tempo zmian częstości alleli w ramach selekcji. Szkodliwe allele recesywne mogą utrzymywać się w populacji przy niskich częstotliwościach, przy czym większość kopii jest przenoszona w heterozygotach, bez żadnych kosztów dla tych osobników. Te rzadkie choroby recesywne są podstawą wielu dziedzicznych zaburzeń genetycznych .
Nomenklatura
W genetyce symbole zaczęły się jako algebraiczne symbole zastępcze. Kiedy jeden allel dominuje nad innym, najstarszą konwencją jest symbolizowanie dominującego allelu wielką literą. Allelowi recesywnemu przypisuje się tę samą literę małą literą. W przykładzie grochu, gdy znana jest relacja dominacji między dwoma allelami, możliwe jest wyznaczenie dominującego allelu, który tworzy okrągły kształt, przez wielką literę symbolu R , a allel recesywny, który daje pomarszczony kształt, przez niższy- symbol obudowy r . Genotypy homozygotyczne dominujące, heterozygotyczne i homozygotyczne recesywne są następnie zapisywane RR , Rr i rr . Możliwe byłoby również oznaczenie tych dwóch alleli jako W i w oraz trzech genotypów WW , Ww i ww , z których dwa pierwsze dawały okrągły groszek, a trzeci pomarszczony. Wybór „ R ” lub „ W ” jako symbolu dominującego allelu nie przesądza o tym, czy allel powodujący fenotyp „okrągły” czy „pomarszczony” u homozygoty jest dominujący.
Gen może mieć kilka alleli. Każdy allel jest symbolizowany przez symbol locus, po którym następuje unikalny indeks górny. U wielu gatunków najczęstszym allelem w dzikiej populacji jest allel typu dzikiego. Jest symbolizowany znakiem + jako indeksem górnym. Inne allele są dominujące lub recesywne w stosunku do allelu typu dzikiego. W przypadku alleli recesywnych symbol locus jest pisany małymi literami. W przypadku alleli o dowolnym stopniu dominacji w stosunku do allelu typu dzikiego, pierwsza litera symbolu locus jest pisana wielką literą. Ay , oto niektóre allele w locus myszy laboratoryjnej, Mus musculus : , dominujący żółty; a + , typ dziki; i bt , czarny podpalany. Allel a bt jest recesywny w stosunku do allelu typu dzikiego, a allel Ay jest kodominujący w stosunku do allelu typu dzikiego. Allel Ay . jest również kodominujący w stosunku do allelu a bt , ale wykazanie tego związku wykracza poza granice zasad nomenklatury genetycznej myszy
Zasady nomenklatury genetycznej ewoluowały, gdy genetyka stała się bardziej złożona. Komitety ustandaryzowały zasady dla niektórych gatunków, ale nie dla wszystkich. Zasady dla jednego gatunku mogą się nieco różnić od zasad dla innego gatunku.
Związek z innymi koncepcjami genetycznymi
Wiele alleli
Chociaż każdy osobnik organizmu diploidalnego ma co najwyżej dwa różne allele w dowolnym locus (z wyjątkiem aneuploidii ), większość genów występuje w dużej liczbie wersji allelicznych w całej populacji. Jeśli allele mają różny wpływ na fenotyp, czasami ich relacje dominacji można opisać jako serię.
Na przykład kolor sierści kotów domowych zależy od szeregu alleli genu TYR (który koduje enzym tyrozynazę ). Allele C , cb ) wytwarzają różne , cs poziomy i ca . (odpowiednio pełny kolor, birmański , syjamski i albinos pigmentu, a tym samym różne poziomy rozcieńczenia koloru Allel C (pełny kolor) jest całkowicie dominujący nad trzema ostatnimi, a allel c a (albinos) jest całkowicie recesywny w stosunku do pierwszych trzech.
autosomalna a związana z płcią
U ludzi i innych gatunków ssaków płeć jest determinowana przez dwa chromosomy płci zwane chromosomem X i chromosomem Y. Ludzkie kobiety są XX ; mężczyźni są XY . Pozostałe pary chromosomów występują u obu płci i nazywane są autosomami ; cechy genetyczne związane z loci na tych chromosomach są opisywane jako autosomalne i mogą być dominujące lub recesywne. Cechy genetyczne na X i Y nazywane są sprzężonymi z płcią, ponieważ są połączone z chromosomami płciowymi, a nie dlatego, że są charakterystyczne dla jednej lub drugiej płci. W praktyce termin ten prawie zawsze odnosi się do X , a na wiele takich cech (takich jak upośledzenie widzenia barwy czerwono-zielonej) nie ma wpływu płeć. Kobiety mają dwie kopie każdego locus genu znalezionego na chromosomie X, tak jak w przypadku autosomów, i obowiązują te same relacje dominacji. Jednak mężczyźni mają tylko jedną kopię każdego locus genu chromosomu X i są opisani jako hemizygotyczni dla tych genów. Chromosom Y jest znacznie mniejszy niż X i zawiera znacznie mniejszy zestaw genów, w tym między innymi te, które wpływają na „męskość”, takie jak gen SRY dla czynnika determinującego jądra . Reguły dominacji dla loci genów sprzężonych z płcią są określone przez ich zachowanie u kobiet: ponieważ samiec ma tylko jeden allel (z wyjątkiem niektórych typów aneuploidii chromosomu Y), allel ten jest zawsze wyrażany niezależnie od tego, czy jest dominujący, czy recesywny. Ptaki mają chromosomy płci przeciwnej: samce ptaków mają chromosomy ZZ, a samice ZW. Jednak dziedziczenie cech przypomina inaczej system XY; Samce zięb zebry mogą przenosić gen białego koloru w swoim jednym z dwóch chromosomów Z, ale samice zawsze mają biały kolor. Koniki polne mają system XO. Kobiety mają XX, ale mężczyźni tylko X. W ogóle nie ma chromosomu Y.
Epistaza
Epistaza [" epi + zastój = siedzieć na wierzchu"] to interakcja między allelami w dwóch różnych loci genów, które wpływają na pojedynczą cechę, co może czasami przypominać interakcję dominacji między dwoma różnymi allelami w tym samym locus. Epistaza modyfikuje charakterystyczny 9:3:3:1 oczekiwany dla dwóch genów nieepistatycznych. Dla dwóch loci rozpoznaje się 14 klas oddziaływań epistatycznych. Jako przykład epistazy recesywnej , jeden locus genu może określać, czy pigment kwiatowy jest żółty ( AA lub Aa ) czy zielony ( aa ), podczas gdy inny locus określa, czy pigment jest wytwarzany ( BB lub Bb ), czy nie ( bb ). W bb kwiaty będą białe, niezależnie od genotypu innego locus jak AA , Aa lub aa . Kombinacja bb nie jest dominująca względem allelu A : raczej gen B wykazuje recesywną epistazę względem genu A , ponieważ locus B , gdy jest homozygotyczny pod względem allelu recesywnego ( bb ), hamuje fenotypową ekspresję locus A. W krzyżówce dwóch roślin AaBb daje to charakterystyczny stosunek 9:3:4 , w tym przypadku żółte : zielone : białe kwiaty.
W epistazie dominującej jeden locus genu może determinować żółty lub zielony pigment, jak w poprzednim przykładzie: AA i Aa są żółte, a aa są zielone. Drugi locus określa, czy prekursor pigmentu jest wytwarzany ( dd ), czy nie ( DD lub Dd ). Tutaj, w DD lub Dd , kwiaty będą bezbarwne niezależnie od genotypu w locus A , z powodu efektu epistatycznego dominującego allelu D. Tak więc w krzyżówce między dwiema roślinami AaDd 3/4 roślin będzie bezbarwnych, a żółty i zielony fenotyp występuje tylko w roślinach dd . Daje to charakterystyczny stosunek 12:3:1 białych : żółtych : zielonych roślin.
Dodatkowa epistaza występuje, gdy dwa loci wpływają na ten sam fenotyp. Na przykład, jeśli kolor pigmentu jest wytwarzany przez CC lub Cc , ale nie cc , oraz przez DD lub Dd , ale nie dd , to pigment nie jest wytwarzany w żadnej kombinacji genotypowej z cc lub dd . Oznacza to, że oba loci muszą mieć co najmniej jeden dominujący allel, aby wytworzyć fenotyp. Daje to charakterystyczny 9: 7 roślin pigmentowanych do niepigmentowanych. W przeciwieństwie do epistazy komplementarnej powstaje roślina bez pigmentu wtedy i tylko wtedy, gdy genotyp to cc i dd , a charakterystyczny stosunek między roślinami pigmentowanymi i niepigmentowanymi wynosi 15: 1 .
Genetyka klasyczna rozważała interakcje epistatyczne między dwoma genami jednocześnie. Obecnie na podstawie genetyki molekularnej jest oczywiste, że wszystkie loci genów są zaangażowane w złożone interakcje z wieloma innymi genami (np. .
Zasada Hardy'ego-Weinberga (oszacowanie częstotliwości nośnej)
Częstotliwość stanu heterozygotycznego (który jest stanem nosiciela cechy recesywnej) można oszacować za pomocą wzoru Hardy'ego-Weinberga :

Ten wzór odnosi się do genu z dokładnie dwoma allelami i wiąże częstość tych alleli w dużej populacji z częstością ich trzech genotypów w tej populacji.
Na przykład, jeśli p jest częstością allelu A , a q jest częstością allelu a , to terminy p2 , 2 pq i q2 są odpowiednio częstością genotypów AA , Aa i aa . Ponieważ gen ma tylko dwa allele, wszystkie allele muszą być A lub a i p + q = 1 . Teraz, jeśli A jest całkowicie dominujący wobec a , to częstości genotypu nosiciela Aa nie można bezpośrednio zaobserwować (ponieważ ma on te same cechy, co homozygotyczny genotyp AA ), jednak można to oszacować na podstawie częstości cechy recesywnej w populacji , ponieważ jest to to samo, co dla homozygotycznego genotypu aa . tzn. można oszacować częstości poszczególnych alleli: q = √ f (aa) , p = 1 − q , az nich można wyprowadzić częstość genotypu nosiciela: f (Aa) = 2 pq .
Formuła ta opiera się na szeregu założeń i dokładnym oszacowaniu częstości występowania cechy recesywnej. Ogólnie rzecz biorąc, każda rzeczywista sytuacja będzie do pewnego stopnia odbiegać od tych założeń, wprowadzając odpowiednie niedokładności do oszacowania. Jeśli cecha recesywna jest rzadka, trudno będzie dokładnie oszacować jej częstość, ponieważ potrzebna będzie bardzo duża próba.
Dominujący kontra korzystny
Właściwość „dominującego” jest czasami mylona z pojęciem korzystnego, a właściwość „recesywnego” jest czasami mylona z pojęciem szkodliwego, ale zjawiska te są różne. Dominacja opisuje fenotyp heterozygot w odniesieniu do fenotypów homozygot i bez względu na stopień, w jakim różne fenotypy mogą być korzystne lub szkodliwe. Ponieważ wiele alleli chorób genetycznych jest recesywnych, a słowo dominacja ma pozytywne konotacje, często przyjmuje się założenie, że dominujący fenotyp jest lepszy pod względem sprawności. Nie jest to jednak zapewnione; jak omówiono poniżej, podczas gdy większość alleli chorób genetycznych jest szkodliwa i recesywna, nie wszystkie choroby genetyczne są recesywne.
Niemniej jednak to zamieszanie było wszechobecne w całej historii genetyki i utrzymuje się do dziś. Rozwiązanie tego zamieszania było jedną z głównych motywacji do opublikowania zasady Hardy'ego-Weinberga .
Mechanizmy molekularne
Molekularne podstawy dominacji były nieznane Mendelowi. Obecnie wiadomo, że locus genu obejmuje długą serię (setki do tysięcy) zasad lub nukleotydów kwasu dezoksyrybonukleinowego (DNA) w określonym punkcie chromosomu. Centralny dogmat biologii molekularnej głosi, że „ DNA sprawia, że RNA tworzy białko ”, to znaczy, że DNA jest transkrybowane w celu utworzenia kopii RNA, a RNA ulega translacji w celu wytworzenia białka. W tym procesie różne allele w locus mogą być transkrybowane lub nie, a jeśli są transkrybowane, mogą być tłumaczone na nieco inne wersje tego samego białka (zwane izoformami ). Białka często działają jako enzymy , które katalizują reakcje chemiczne w komórce, które bezpośrednio lub pośrednio wytwarzają fenotypy. Mutacje w genomie mogą zmieniać aktywność katalityczną, a tym samym wpływać na dominację. W każdym organizmie diploidalnym sekwencje DNA dwóch alleli obecnych w dowolnym locus genu mogą być identyczne (homozygotyczne) lub różne (heterozygotyczne). Nawet jeśli locus genu jest heterozygotyczny na poziomie sekwencji DNA, białka wytwarzane przez każdy allel mogą być identyczne. W przypadku braku jakiejkolwiek różnicy między produktami białkowymi, żaden allel nie może być uznany za dominujący (patrz współdominacja powyżej). Nawet jeśli te dwa produkty białkowe różnią się nieco ( alozymy ), jest prawdopodobne, że wytwarzają ten sam fenotyp pod względem działania enzymów i ponownie nie można powiedzieć, że żaden allel nie jest dominujący.
Zygotyczność
Historycznie rzecz biorąc, prawo Mendla o niezależnym asortymencie zakładało, że allele będą sortować niezależnie, przy czym jeden allel jest „dominujący”. Zygotyczność , stopień podobieństwa alleli organizmu, może wpływać na dominację. W organizmie diploidalnym byłyby one zdefiniowane przez haplotypów alleli. Haploidia genów może skutkować pojedynczym, funkcjonalnym allelem wytwarzającym wystarczającą ilość białka do wytworzenia fenotypu identycznego z fenotypem homozygoty . Możliwe są trzy ogólne typy interakcji haplotypów:
- Haplosuficjencja. W diploidzie funkcjonalny allel genu haplosufitowego byłby uważany za dominujący, podczas gdy niefunkcjonalny allel byłby uważany za recesywny. Załóżmy na przykład, że standardowa ilość enzymu wytwarzanego w funkcjonalnej homozygocie wynosi 100%, przy czym każdy z dwóch funkcjonalnych alleli ma po 50%. Pojedynczy funkcjonalny allel w heterozygocie wytwarza 50% standardowej ilości enzymu, która jest wystarczająca do wytworzenia standardowego fenotypu. Jeśli heterozygota i homozygota z allelem funkcjonalnym mają identyczne fenotypy, allel funkcjonalny dominuje nad allelem niefunkcjonalnym. Dzieje się tak w locus genu albinosa: heterozygota wytwarza wystarczającą ilość enzymu do przekształcenia prekursora pigmentu w melaninę, a osobnik ma standardową pigmentację. Na przykład u ludzi i innych organizmów skóra bez pigmentu o albinosów powstaje, gdy osobnik jest homozygotyczny pod względem allelu kodującego niefunkcjonalną wersję enzymu potrzebnego do produkcji melaniny będącej pigmentem skóry .
- Niekompletna haploinsuficiency. Rzadziej obecność pojedynczego funkcjonalnego allelu daje fenotyp, który nie jest normalny, ale mniej poważny niż fenotyp niefunkcjonalnej homozygoty. Dzieje się tak, gdy funkcjonalny allel nie jest haplo-wystarczający, dlatego w tych przypadkach zwykle stosuje się terminy haplo-niedobór i niepełna dominacja. Interakcja pośrednia występuje, gdy genotyp heterozygotyczny wytwarza fenotyp pośredni między dwiema homozygotami. W zależności od tego, którą z dwóch homozygot najbardziej przypomina heterozygota, mówi się, że jeden allel wykazuje niepełną dominację nad drugim. Na przykład u ludzi Hb jest odpowiedzialne za białko łańcucha beta ( HBB ), które jest jednym z dwóch białek globiny tworzących hemoglobinę jako barwnik krwi . Wiele osób jest homozygotami pod względem allelu zwanego Hb A ; niektóre osoby są nosicielami alternatywnego allelu zwanego Hb S , jako homozygoty lub heterozygoty. Cząsteczki hemoglobiny Hb S / Hb S ulegają zmianie kształtu, która zniekształca morfologię krwinek czerwonych i powoduje ciężką, zagrażającą życiu postać niedokrwistości zwaną niedokrwistością sierpowatokrwinkową . Osoby heterozygotyczne pod względem Hb A / Hb S pod względem tego allelu mają znacznie łagodniejszą postać niedokrwistości zwaną niedokrwistością sierpowatokrwinkową . Ponieważ fenotyp choroby Hb A / Hb S jest bardziej podobny, ale nie identyczny z homozygotą Hb A / Hb A , mówi się, że allel Hb A jest niecałkowicie dominujący w stosunku do allelu Hb S.
- Całkowita haploinsuficjencja. Pojedynczy funkcjonalny allel w heterozygocie może wytwarzać niewystarczający produkt genu dla jakiejkolwiek funkcji genu, powodując, że zwykle niefunkcjonalne allele stają się dominujące. Fenotyp będzie wtedy przypominał fenotyp homozygoty z niefunkcjonalnym allelem zamiast typu dzikiego. Można by powiedzieć, że niefunkcjonalny allel jest dominujący w stosunku do funkcjonalnego allelu fenotypu typu dzikiego. Taka sytuacja może wystąpić, gdy niefunkcjonalny allel wytwarza wadliwe białko, które zakłóca prawidłowe funkcjonowanie białka wytwarzanego przez standardowy allel. Obecność wadliwego białka „dominuje” w białku standardowym, a fenotyp choroby heterozygoty bardziej przypomina fenotyp homozygoty pod względem dwóch wadliwych alleli. Termin „dominujący” jest często błędnie stosowany do wadliwych alleli, których homozygotyczny fenotyp nie został zbadany, ale które powodują odrębny fenotyp, gdy są heterozygotyczne z normalnym allelem. Zjawisko to występuje w wielu z powtórzeniami trinukleotydów , jednym z przykładów jest choroba Huntingtona . W chorobie Huntingtona całkowita haploinsuficjencja powoduje dominujący efekt zmutowanego białka. Zwykle osoba ma około 20 powtórzeń nukleotydów CAG w swoim genie HTT, ale osoby z chorobą Huntingtona mają ponad 40 powtórzeń CAG. Innym przykładem jest zespół Marfana, dziedziczna choroba tkanki łącznej, spowodowana mutacją w genie fibryliny-1 (FBN1). Jedna normalna kopia genu FBN1 jest dziedziczona od jednego rodzica, podczas gdy dominująca nieprawidłowa kopia genu FBN1 jest dziedziczona przez innego rodzica.
Mutacje dominujące negatywne
Wiele białek jest zwykle aktywnych w postaci multimeru, agregatu wielu kopii tego samego białka, inaczej znanego jako białko homomultimeryczne lub białko homooligomeryczne . W rzeczywistości większość z 83 000 różnych enzymów z 9800 różnych organizmów w bazie danych enzymów BRENDA to homooligomery. Kiedy wersja białka typu dzikiego jest obecna wraz z wersją zmutowaną, może powstać mieszany multimer. Mutacja, która prowadzi do zmutowanego białka, które zakłóca aktywność białka typu dzikiego w multimerze, jest mutacją dominującą negatywną.
Mutacja dominująca negatywna może powstać w ludzkiej komórce somatycznej i zapewnić przewagę proliferacyjną zmutowanej komórce, prowadząc do jej ekspansji klonalnej. Na przykład dominująca negatywna mutacja w genie niezbędnym do normalnego procesu zaprogramowanej śmierci komórki ( apoptozy ) w odpowiedzi na uszkodzenie DNA może uczynić komórkę odporną na apoptozę. Umożliwi to namnażanie się klonu nawet przy nadmiernym uszkodzeniu DNA. Takie mutacje dominujące-ujemne występują w genie supresorowym guza p53 . Białko P53 typu dzikiego jest normalnie obecne jako multimer czterech białek (oligotetramer). Dominujące negatywne p53 występują w wielu różnych typach raka i zmianach przedrakowych (np. guzach mózgu, raku piersi, zmianach przedrakowych jamy ustnej i raku jamy ustnej).
Mutacje dominujące negatywne występują również w innych genach supresorowych nowotworów. Na przykład zidentyfikowano dwie dominujące negatywne mutacje linii zarodkowej w zmutowanym genie Ataxia telangiectasia (ATM), który zwiększa podatność na raka piersi. Dominujące negatywne mutacje czynnika transkrypcyjnego C/EBPα mogą powodować ostrą białaczkę szpikową. Odziedziczone dominujące mutacje negatywne mogą również zwiększać ryzyko chorób innych niż rak. Dominujące negatywne mutacje w receptorze gamma aktywowanym przez proliferatory peroksysomów (PPARγ) są związane z ciężką insulinoopornością, cukrzycą i nadciśnieniem.
Mutacje dominujące negatywne opisano również w organizmach innych niż ludzie. W rzeczywistości pierwsze badanie opisujące zmutowane białko hamujące normalną funkcję białka typu dzikiego w mieszanym multimerze dotyczyło GP37 białka włókna ogona T4 bakteriofaga. Wydaje się, że mutacje, które wytwarzają raczej skrócone białko niż zmutowane białko pełnej długości, mają najsilniejszy dominujący negatywny wpływ w badaniach P53, ATM, C / EBPα i bakteriofaga T4 GP37.

Dziedziczne defekty enzymów są na ogół dziedziczone w sposób autosomalny, ponieważ jest więcej chromosomów innych niż chromosomy X niż chromosomy X, oraz w sposób recesywny, ponieważ enzymy z nienaruszonych genów są na ogół wystarczające do zapobiegania objawom u nosicieli. Wyjątki obejmują przypadki haploinsufficiency , w których nienaruszony gen nie może zrekompensować dotkniętego genu.
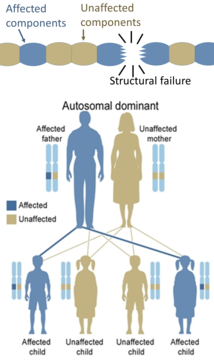
Z drugiej strony, dziedziczne defekty białek strukturalnych (takie jak osteogenesis imperfecta , zespół Marfana i zespoły Ehlersa-Danlosa ) są na ogół autosomalne dominujące, ponieważ wystarczy, że niektóre elementy są wadliwe, aby cała struktura była dysfunkcyjna. Jest to proces dominujący-negatywny, w którym zmutowany produkt genu niekorzystnie wpływa na niezmutowany produkt genu w tej samej komórce.


Dominujące i recesywne choroby genetyczne u ludzi
U ludzi wiele cech genetycznych lub chorób jest po prostu klasyfikowanych jako „dominujące” lub „recesywne”. Zwłaszcza w przypadku tak zwanych chorób recesywnych, które rzeczywiście są czynnikiem genów recesywnych, ale mogą nadmiernie upraszczać podstawowe podstawy molekularne i prowadzić do niezrozumienia natury dominacji.
Na przykład recesywna choroba genetyczna, fenyloketonuria (PKU), wynika z dowolnego z dużej liczby (>60) alleli w locus genu enzymu hydroksylazy fenyloalaniny ( WWA ). Wiele z tych alleli wytwarza niewiele WWA lub nie wytwarza ich wcale , w wyniku czego substrat fenyloalaniny (Phe) i jej metaboliczne produkty uboczne gromadzą się w ośrodkowym układzie nerwowym i mogą powodować poważną niepełnosprawność intelektualną, jeśli nie są leczone.
Aby zilustrować te niuanse, w poniższej tabeli przedstawiono genotypy i fenotypowe konsekwencje interakcji między trzema hipotetycznymi allelami WWA:
| Genotyp | działalność PAH | [ Phe ] stęż | PKU? |
|---|---|---|---|
| AA | 100% | 60 μM | NIE |
| AB | 30% | 120 μM | NIE |
| CC | 5% | 200 ~ 300 μM | Hiperfenyloalaninemia |
| nocleg ze śniadaniem | 0,3% | 600 ~ 2400 μM | Tak |
U zdrowych osób, homozygotycznych pod względem standardowego funkcjonalnego allelu ( AA ), aktywność WWA jest standardowa (100%), a stężenie fenyloalaniny we krwi [ Phe ] wynosi około 60 μM (= μmol/L ). U nieleczonych osób homozygotycznych pod względem jednego z alleli PKU ( BB ) aktywność WWA jest bliska zeru, [Phe] od dziesięciu do czterdziestu razy standardowa, a u danej osoby manifestuje się PKU.
U heterozygoty AB aktywność PAH wynosi tylko 30% (a nie 50%) normy, [ Phe ] we krwi jest dwukrotnie podwyższone, a osoba nie manifestuje PKU. Zatem A dominuje nad allelem B pod względem PKU, ale allel B jest niecałkowicie dominujący nad allelem A pod względem efektu molekularnego, określenie poziomu aktywności WWA (0,3% < 30% << 100%) . Wreszcie A jest niecałkowicie dominujący w stosunku do allelu B w odniesieniu do [Phe], ponieważ 60 μM <120 μM << 600 μM. Zauważmy jeszcze raz, że dla kwestii dominacji nie ma znaczenia to, że allel recesywny wytwarza bardziej ekstremalny fenotyp [Phe].
W przypadku trzeciego allelu C homozygota CC wytwarza bardzo małą ilość enzymu WWA , co powoduje nieco podwyższony poziom [ Phe ] we krwi, stan zwany hiperfenyloalaninemią , który nie powoduje niepełnosprawności intelektualnej.
Oznacza to, że relacje dominacji dowolnych dwóch alleli mogą się różnić w zależności od tego, który aspekt fenotypu jest brany pod uwagę. Zwykle bardziej przydatne jest mówienie o fenotypowych konsekwencjach interakcji allelicznych zaangażowanych w dowolny genotyp, niż próba wtłoczenia ich w kategorie dominujące i recesywne.
Zobacz też
- „Notatki on-line do biologii 2250 - Zasady genetyki” . Uniwersytet Pamięci Nowej Fundlandii.
- Internetowe dziedziczenie mendlowskie u człowieka (OMIM): Hemoglobina — Beta Locus; HBB - 141900 — Anemia sierpowata
- Internetowe dziedziczenie mendlowskie u człowieka (OMIM): glikozylotransferaza ABO - 110300 — grupy krwi ABO