Kompleks kinazy cyklinozależnej
Kompleks kinazy cyklinozależnej ( CDKC , cyklina-CDK ) jest kompleksem białkowym utworzonym przez połączenie nieaktywnej katalitycznej podjednostki kinazy białkowej, kinazy zależnej od cykliny (CDK), z podjednostką regulatorową, cykliną . Gdy kinazy cyklinozależne zwiążą się z cykliną, utworzony kompleks jest w stanie aktywowanym. Specyficzność substratowa aktywowanego kompleksu jest ustalana głównie przez związaną z nim cyklinę. Aktywność CDKC jest kontrolowana przez fosforylację białek docelowych, jak również wiązanie białek hamujących.
Struktura i regulacja

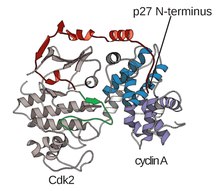
Struktura CDK w kompleksie z podjednostkami cykliny (CDKC) była od dawna celem biologów strukturalnych i komórkowych, począwszy od lat 90. XX wieku, kiedy Brown i in. oraz w tym samym roku Jeffery i in. rozwiązał strukturę kompleksu ludzkiej cykliny A-CDK2 z rozdzielczością 2,3 angstremów. Od tego czasu określono, że wiele struktur CDK ma wyższą rozdzielczość, w tym struktury CDK2 i CDK2 związane z różnymi substratami, jak widać na rycinie 1. Struktury o wysokiej rozdzielczości istnieją łącznie dla około 25 kompleksów CDK-cyklina w białku Bank danych . W zależności od funkcji istnieją dwie ogólne populacje złożonych struktur CDK-cyklina, forma otwarta i zamknięta. Różnica między postaciami polega na wiązaniu partnerów cykliny, gdzie zamknięte kompleksy postaci wiążą CDK-cyklina zarówno na C, jak i na N-końcu pętli aktywacyjnej CDK, podczas gdy partnerzy w postaci otwartej wiążą się tylko na N-końcu. Struktury w formie otwartej odpowiadają najczęściej kompleksom zaangażowanym w regulację transkrypcji (CDK 8, 9, 12 i 13), podczas gdy forma zamknięta kompleksu CDK-cyklina jest najczęściej zaangażowana w progresję i regulację cyklu komórkowego (CDK 1, 2, 6) . Te odrębne role nie różnią się jednak znacząco pod względem homologii sekwencji między składnikami CDK. W szczególności, wśród tych znanych struktur wydają się istnieć cztery główne regiony konserwatywne: N-końcowa pętla bogata w glicynę, region zawiasowy, helisa αC i miejsce regulacji pętli T.
Pętla aktywacji
Pętla aktywacyjna , określana również jako pętla T, jest regionem CDK (między motywami DFG i APE w wielu CDK), który jest aktywny enzymatycznie, gdy CDK jest związany ze swoim specyficznym dla funkcji partnerem. W kompleksach CDK-cyklina ten region aktywacji składa się z konserwowanej helisy αL-12 i zawiera kluczową resztę ulegającą fosforylacji (zwykle treoninę dla partnerów cykliny CDK, ale obejmuje również serynę i tyrozynę), która pośredniczy w aktywności enzymatycznej CDK. To właśnie przy tej istotnej reszcie (T160 w kompleksach CDK2, T177 w kompleksach CDK6) zachodzi enzymatyczna fosforylacja ATP kompleksów CDK-cyklina przez CAK (kinaza aktywująca cykliny, odnosząca się do kompleksu CDK7-cyklina H w komórkach ludzkich). Po hydrolizie ATP do fosforylacji w tym miejscu, kompleksy te są w stanie wypełnić swoją zamierzoną funkcję, fosforylację celów komórkowych. Należy zauważyć, że w CDK 1, 2 i 6 pętla T i oddzielny region C-końcowy są głównymi miejscami wiązania cyklin w CDK, a to, które cykliny są związane z każdą z tych CDK, odbywa się za pośrednictwem określoną sekwencję pętli T miejsca aktywacji. Te miejsca wiązania cyklin są regionami o największej zmienności w CDK pomimo stosunkowo wysokiej homologii sekwencji otaczającej motyw helisy αL-12 tego składnika strukturalnego.
Region bogaty w glicynę
Pętla bogata w glicynę (pętla bogata w Gly), jak widać w resztach 12-16 w CDK2, koduje konserwatywny motyw GXGXXG zarówno w modelach drożdżowych, jak i zwierzęcych. Region regulatorowy podlega zróżnicowanej fosforylacji na resztach innych niż glicyna w tym motywie, co powoduje, że to miejsce podlega fosforylacji kinazy hamującej Wee1 i / lub Myt1 oraz defosforylacji Cdc25 u ssaków. Ta odwracalna fosforylacja w pętli bogatej w Gly w CDK2 występuje w Y15, gdzie aktywność była dalej badana. Badanie tej reszty wykazało, że fosforylacja promuje zmianę konformacyjną, która zapobiega wiązaniu ATP i substratu przez steryczną interferencję z tymi niezbędnymi miejscami wiązania w pętli aktywacyjnej kompleksów CDK-cyklina. Ta aktywność jest wspomagana przez godną uwagi elastyczność, jaką bogata w Gly pętla ma w strukturze większości CDK, co pozwala na jej obrót w kierunku pętli aktywacyjnej, aby mieć znaczący wpływ na zmniejszenie powinowactwa substratu bez większych zmian w ogólnej strukturze kompleksu CDK-cyklina.
Region zawiasów
Konserwatywny region zawiasowy CDK w komórkach eukariotycznych działa jako niezbędny pomost między pętlą bogatą w Gly a pętlą aktywacyjną. CDK charakteryzują się N-końcowym płatem, który jest głównie skręconą beta-kartką połączoną przez ten region zawiasowy z dominującym w helisie alfa płatem C-końcowym. Omawiając pętlę T i pętlę bogatą w Gly, należy zauważyć, że te regiony, które muszą być w stanie oddziaływać przestrzennie, aby wykonywać swoje funkcje biochemiczne, leżą na przeciwległych płatach samego CDK. Tak więc ten region zawiasowy, którego długość może się nieznacznie różnić między typem CDK a kompleksem CDK-cyklina, łączy istotne regiony regulatorowe CDK poprzez połączenie tych płatów i odgrywa kluczową rolę w powstałej strukturze kompleksów CDK-cyklina poprzez odpowiednie ukierunkowanie ATP dla łatwej katalizy reakcji fosforylacji przez zmontowany kompleks.
helisa αC
Region αC-Helix jest wysoce konserwatywny w wielu kinomach ssaków (rodzina kinaz ). Jego głównym zadaniem jest utrzymanie allosterycznej kontroli nad miejscem aktywnym kinazy. Ta kontrola manifestuje się w kompleksach CDK-cyklina przez swoiste zapobieganie aktywności CDK, dopóki nie zwiąże się ze swoim partnerem regulatorowym (tj. cykliną lub innym białkiem partnerskim). To wiązanie powoduje zmianę konformacyjną w regionie αC-Helix CDK i pozwala na przeniesienie go z szczeliny miejsca aktywnego i kończy początkowy proces aktywacji pętli T. Biorąc pod uwagę, że region ten jest tak konserwatywny w całej nadrodzinie białkowej kinaz, ten mechanizm, w którym wykazano, że helisa αC wygina się z N-końcowego płata kinazy, umożliwiając zwiększony dostęp do helisy αL-12, która leży w pętli T, jest uważany za potencjalny cel dla rozwoju leków.
Cykl komórkowy
Cykl komórkowy drożdży
Chociaż kompleksy te mają różnorodne funkcje, CDKC są najbardziej znane ze swojej roli w cyklu komórkowym . Początkowo badania prowadzono na Schizosaccharomyces pombe i Saccharomyces cerevisiae (drożdże). S. pombe i S. cerevisiae są najbardziej znane ze swojego związku odpowiednio z pojedynczym Cdk, Cdc2 i Cdc28, które tworzą kompleksy z kilkoma różnymi cyklinami. W zależności od cykliny wpływa to na różne części cyklu komórkowego. Na przykład u S. pombe Cdc2 łączy się z Cdk13, tworząc kompleks Cdk13-Cdc2. W S. cerevisiae asocjacja Cdc28 z cyklinami Cln1, Cln2 lub Cln3 powoduje przejście z fazy G1 do fazy S. W fazie S Cln1 i Cln2 dysocjują z Cdc28 i tworzą się kompleksy między Cdc28 i Clb5 lub Clb6. W fazie G2 kompleksy fazy G2 utworzone z połączenia między Cdc28 i Clb1, Clb2, Clb3 lub Clb4 powodują przejście od do fazy M (mitotycznej). Kompleksy te są również obecne we wczesnej fazie M. Zobacz tabelę 1, aby zapoznać się z podsumowaniem CDKC drożdży.
- Tabela 1. CDKC związane z fazami cyklu komórkowego w drożdżach
| CDK | cyklina | Faza cyklu komórkowego |
|---|---|---|
| Cdc2 ( S. pombe ) | Cdc13 | przejście fazowe G2 do M; wczesna faza M |
| Cdc28 ( S. cerevisiae ) | Cln1, Cln2 | Przejście fazowe G1 do S |
| Cdc28 | Clb5, Clb6 | Faza S |
| Cdc28 | CLB1, CLB2, CLB3, CLB4 | przejście fazowe G2 do M; wczesna faza M |
Z tego, co wiadomo na temat kompleksów powstających podczas każdej fazy cyklu komórkowego w drożdżach, wyłoniły się proponowane modele oparte na ważnych miejscach fosforylacji i zaangażowanych czynnikach transkrypcyjnych.
Cykl komórkowy ssaków
Korzystając z informacji uzyskanych dzięki badaniom cyklu komórkowego drożdży, poczyniono znaczne postępy w zakresie cyklu komórkowego ssaków. Ustalono, że cykle komórkowe są podobne, a CDKC bezpośrednio lub pośrednio wpływają na przebieg cyklu komórkowego. Jak wspomniano wcześniej, w drożdżach tylko jedna kinaza zależna od cyklin (CDK) jest związana z kilkoma różnymi cyklinami. Jednak w komórkach ssaków kilka różnych CDK wiąże się z różnymi cyklinami, tworząc CDKC. jako ludzki Cdc2), pierwszy zidentyfikowany ludzki CDK, łączy się z cyklinami A lub B. Kompleksy CyclinA/B-Cdk1 kierują przejściem między fazą G2 a fazą M, a także wczesną fazą M. Inny ssaczy CDK, Cdk2, może tworzyć kompleksy z cyklinami D1, D2, D3, E lub A. Cdk4 i Cdk6 oddziałują z cyklinami D1, D2 i D3. Badania wykazały, że nie ma różnicy między cyklinami CDKC D1-Cdk4/6, dlatego wszelkie unikalne właściwości można prawdopodobnie powiązać ze specyficznością substratową lub aktywacją. Podczas gdy poziomy CDK pozostają dość stałe przez cały cykl komórkowy, poziomy cyklin ulegają wahaniom. Fluktuacja kontroluje aktywację kompleksów cyklina-CDK i ostatecznie postęp w całym cyklu. Patrz Tabela 2, aby zapoznać się z podsumowaniem CDKC komórek ssaków zaangażowanych w cykl komórkowy.
- Tabela 2. CDKC związane z fazami cyklu komórkowego w komórkach ssaków
| CDK | cyklina | Faza cyklu komórkowego | Niecykliczne białka partnerskie |
|---|---|---|---|
| Cdk1 (Cdc2) | Cykliny A i B | przejście fazowe G2 do M; wczesna faza M | Cks1 i Cks2 |
| cdk2 | Cykliny D1, D2, D3 | Faza G1 | KAP, Cks1, p27KIP1 i Spy-1 |
| cdk2 | Cyklina E | Przejście fazowe G1 do S | KAP, Cks1, p27KIP1 i Spy-1 |
| cdk2 | Cyklina A | Faza S | KAP, Cks1, p27KIP1 i Spy-1 |
| cdk4 | Cykliny D1, D2, D3 | Faza G1 | HSP90-Cdc37 |
| cdk6 | Cykliny D1, D2, D3 | Faza G1 | p16INK4A, p19INK4D i P18INK4C-cyklina K |
| cdk8 | Cyklina C | --- | --- |
| cdk9 | Cyklin T | --- | Tat, AFF4 i TAR |
| cdk12 | Cyklin K | --- | --- |
| cdk13 | Cyklin K | --- | --- |

G 1 do S
Podczas późnej fazy G1 CDKC wiążą i fosforylują członków rodziny białek siatkówczaka (Rb) . Członkowie rodziny białek Rb są supresorami nowotworów, które zapobiegają niekontrolowanej proliferacji komórek, która mogłaby wystąpić podczas tworzenia się guza. Jednak uważa się również, że pRbs hamują geny wymagane do przejścia z fazy G1 do fazy S. Kiedy komórka jest gotowa do przejścia do następnej fazy, CDKC, cyklina D1-Cdk4 i cyklina D1-Cdk6 fosforylują pRB, po czym następuje dodatkowa fosforylacja z cykliny E-Cdk2 CDKC. Po wystąpieniu fosforylacji czynniki transkrypcyjne są następnie uwalniane w celu nieodwracalnej inaktywacji pRB i następuje przejście do fazy S cyklu komórkowego. Cyklina E-Cdk2 CDKC utworzona w fazie G1 pomaga następnie w inicjacji replikacji DNA podczas fazy S.
G 2 do M
Pod koniec fazy S cyklina A jest związana z Cdk1 i Cdk2. Podczas fazy G2 cyklina A ulega degradacji, podczas gdy cyklina B jest syntetyzowana i tworzą się kompleksy cykliny B-Cdk1. Kompleksy cykliny B-Cdk1 są nie tylko ważne dla przejścia do fazy M, ale te CDKC odgrywają rolę w następujących procesach regulacyjnych i strukturalnych:
- Kondensacja chromosomów
- Fragmentacja sieci Golgiego
- Rozpad blaszki jądrowej
Inaktywacja kompleksu cykliny B-Cdk1 poprzez degradację cykliny B jest konieczna do wyjścia z fazy M cyklu komórkowego.
Inny
Chociaż większość znanych CDKC bierze udział w cyklu komórkowym, nie wszystkie kompleksy kinazy działają w ten sposób. Badania wykazały, że inne CDKC, takie jak cyklina k-Cdk9 i cyklina T1-Cdk9, biorą udział w stres replikacyjny i wpływają na transkrypcję . Ponadto kompleksy cykliny H-Cdk7 mogą odgrywać rolę w mejozie w męskich komórkach rozrodczych i wykazano, że biorą również udział w czynnościach transkrypcyjnych.