Photobacterium profundum

|
|
| Photobacterium profundum | |
|---|---|
| Obraz konfokalny szczepu P. profundum SS9 wykazującego ekspresję zielonego białka fluorescencyjnego . Pręt: 10μm | |
| Klasyfikacja naukowa | |
| Domena: | Bakteria |
| Gromada: | pseudomonadota |
| Klasa: | Gammaproteobakterie |
| Zamówienie: | Wibracje |
| Rodzina: | Vibrionaceae |
| Rodzaj: | Fotobakterie |
| Gatunek: |
P. głęboka
|
| Nazwa dwumianowa | |
|
Photobacterium profundum Nogi i in. 1998
|
|
Photobacterium profundum to głębinowa Gammaproteobacterium , należąca do rodziny Vibrionaceae i rodzaju Photobacterium . Podobnie jak inni członkowie tego rodzaju, P. profundum jest organizmem morskim i ma dwa koliste chromosomy . P. profundum jest Gram-ujemną pałeczką, która może rosnąć w temperaturach od 0°C do 25°C i ciśnieniach od 0,1 MPa do 70 MPa w zależności od szczepu. Ma zapotrzebowanie na sól, jest w stanie metabolizować szeroką gamę węglowodanów prostych i złożonych i ma dwa systemy wici . Komórki mają kształt pręta, długość 2-4 μm i szerokość 0,8-1,0 μm, z pojedynczą wicią bez osłonki. Bakteria ta została pierwotnie wyizolowana w 1986 roku z Morza Sulu i obecnie hoduje się 4 dzikie szczepy P. profundum (szczepy SS9, 3TCK, DJS4 i 1230).
Photobacterium profundum szczep SS9 ma optymalny wzrost w temperaturze 15 ° C i 28 MPa, co czyni go zarówno psychrofilem , jak i piezofilem . P. profundum szczep 3TCK, wyizolowany z Zatoki San Diego , rośnie optymalnie w temperaturze 9°C i 0,1 MPa, a P. profundum szczep DSJ4, wyizolowany z rowu Ryukyu u wybrzeży Japonii na głębokości 5110 m, rośnie optymalnie w temperaturze 10°C i 10 MPa. Na podstawie sekwencji 16S rRNA P. profundum jest blisko spokrewniony z rodzajem Vibrio , z których najważniejszym gatunkiem jest ludzki patogen Vibrio cholerae .
W szczepie SS9 wykazano, że kilka genów odpowiedzi na stres jest regulowanych w górę w odpowiedzi na ciśnienie atmosferyczne, obejmują one htpG, dnaK, dnaJ i groEL. Rodzaje i obfitość łańcuchów kwasów tłuszczowych w błonie komórkowej również reagują na zmiany ciśnienia i temperatury. W niskiej temperaturze i pod wysokim ciśnieniem szczep SS9 zwiększa obfitość jedno- i wielonienasyconych kwasów tłuszczowych. Ma to wpływ na zwiększenie płynności membrany poprzez zmniejszenie upakowania łańcuchów kwasów tłuszczowych, co skutkuje raczej strukturą ciekłokrystaliczną w membranie niż strukturą żelową. Białko błony zewnętrznej Wykazano, że OmpH jest regulowany w górę przy podwyższonym ciśnieniu, odwrotnie jest w przypadku białka błony zewnętrznej OmpL, które jest regulowane w górę w odpowiedzi na niskie ciśnienie.
W 2005 roku Vezzi i in. opublikował sekwencję genomu szczepu P. profundum SS9. Genom P. profundum składa się z kolistego chromosomu 4,1 Mbp, mniejszego kolistego chromosomu 2,2 Mbp, jak również kolistego plazmidu 80 kbp . Szczep SS9 ma 14 genów rybosomalnego RNA (rRNA) na chromosomie 1 i 1 na chromosomie 2; jest to największa liczba genów rRNA znalezionych w jakiejkolwiek bakterii. Chromosom 1 składa się głównie z genów niezbędnych do wzrostu, podczas gdy chromosom 2 wydaje się być dużym plazmidem, który w ewolucyjnej skali czasu zyskał kilka elementów transpozycyjnych . W genomie P. profundum znajduje się duża liczba otwartych ramek odczytu (ORF), które są unikalne dla SS9 i nie występują u innych członków rodziny Vibrionaceae .
Sekwencja genomu uwidoczniła również pełny szlak Sticklanda do fermentacji aminokwasów ; był to pierwszy przypadek zidentyfikowania tej ścieżki w tlenowej . W tej bakterii obecne są również dwie kompletne ścieżki syntazy ATP F1F0 (po jednej na każdym chromosomie): może to wyjaśniać jej zdolność do wytwarzania ATP zarówno pod wysokim, jak i niskim ciśnieniem.
Po tej pracy pojawił się kolejny artykuł w 2005 roku autorstwa Campanaro i in. w którym szczegółowo opisano mikromacierzy porównujące ekspresję genów w suboptymalnych, optymalnych i supraoptymalnych temperaturach i ciśnieniu dla szczepów SS9, 3TCK i DSJ4. Campanaro i in. wykazało, że istnieje 544 rozbieżnych lub brakujących ORF w genomie 3TCK i 562 rozbieżnych lub brakujących ORF w chromosomach DSJ4 w porównaniu z tym z SS9. W artykule tym podkreślono również, że 3TCK nie ma bocznego systemu wici, który jest regulowany w górę w SS9 przy podwyższonym ciśnieniu, jak również brak 3 fagami z 3TCK i 4 regionów związanych z fagami z DSJ4.
Krajobraz transkrypcyjny dzikiego szczepu DB110 i mutanta toxR TW30 zbadano za pomocą sekwencjonowania nowej generacji . ToxR jest przezbłonowym białkiem wiążącym DNA, odkrytym po raz pierwszy w Vibrio cholerae , gdzie reguluje znaczną liczbę genów zaangażowanych w adaptację do środowiska i wirulencję . U P. profundum na obfitość i aktywność tego białka wpływa ciśnienie hydrostatyczne, a jego rola związana jest z regulacją genów w sposób zależny od ciśnienia. Wyniki uzyskane z z RNA-seq ujawniły złożony wzorzec ekspresji z grupą 22 genów o profilach ekspresji podobnych do OmpH, czyli białka błony zewnętrznej transkrybowanego w odpowiedzi na wysokie ciśnienie hydrostatyczne. Co więcej, RNA-seq umożliwiło dogłębną charakterystykę krajobrazu transkrypcyjnego, co doprowadziło do identyfikacji 460 domniemanych małych genów RNA i wykrycia 298 nieznanych wcześniej genów kodujących białka. Przewidywanie w całym genomie operonu , miejsc rozpoczęcia i zakończenia transkrypcji ujawniło nieoczekiwanie dużą liczbę genów (992) z dużymi 5'-UTR, wystarczająco długimi, aby pomieścić cis-regulatorowe struktury RNA, co sugeruje korelację między rozmiarem regionu międzygenowego a długością UTR.
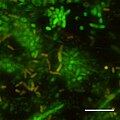
Obraz CSLM początkowych etapów tworzenia biofilmu w Photobacterium profundum SS9 przy użyciu barwników żywych/martwych. Ciśnienie = 28 MPa Temperatura = 15 stopni C Bar = 10 mikrometrów

CSLM Photobacterium profundum SS9 wykazujące ekspresję GFP. Pasek = 10 mikrometrów

