Syfonofory

|
|
| Siphonophorae | |
|---|---|
| (A) Rhizophysa eysenhardtii pasek łuski = 1 cm, (B) Bathyphysa conifera 2 cm, (C) Hippopodius hippopus 5 mm, (D) Kephyes hiulcus 2 mm (E) Desmophyes haematogaster 5 mm (F) Sphaeronectes christiansonae 2 mm, (G) Praya dubia 4 cm (1,6 cala), (H) Apolemia sp. 1 cm, (I) Lychnagalma utricularia 1 cm, (J) Nanomia sp. 1 cm, (K) Physophora hydrostatica 5 mm | |
| Klasyfikacja naukowa | |
| Królestwo: | Animalia |
| Gromada: | Cnidaria |
| Klasa: | hydrozoa |
| Podklasa: | hydroidolina |
| Zamówienie: |
Siphonophorae Eschscholtz , 1829 |
| Podzlecenia | |
| Synonimy | |
|
|
Siphonophorae (z greckiego siphōn „rurka” + pherein „niedźwiedź”) to rząd w obrębie Hydrozoa , który jest klasą organizmów morskich należących do gromady Cnidaria . Według Światowego Rejestru Gatunków Morskich , rząd obejmuje do tej pory 175 gatunków.
Chociaż syfonofor może wydawać się pojedynczym organizmem , każdy okaz jest w rzeczywistości organizmem kolonialnym złożonym z zooidów meduzoidalnych i polipowatych , które są wyspecjalizowane morfologicznie i funkcjonalnie. Zooidy to jednostki wielokomórkowe, które rozwijają się z pojedynczego zapłodnionego jaja i łączą się, tworząc funkcjonalne kolonie zdolne do rozmnażania, trawienia, unoszenia się na wodzie, utrzymywania pozycji ciała i używania napędu odrzutowego do poruszania się. Większość kolonii to długie, cienkie, przezroczyste pływaki żyjące w strefie pelagicznej .
Podobnie jak inne hydrozoany , niektóre syfonofory emitują światło, aby przyciągać i atakować zdobycz. Podczas gdy wiele zwierząt morskich wytwarza niebieską i zieloną bioluminescencję , syfonofor z rodzaju Erenna był dopiero drugą formą życia, która wytwarzała czerwone światło (pierwszą była bezłuskowa ważka Chirostomias pliopterus ).
Anatomia i morfologia
Charakterystyka kolonii
Syfonofory to hydrozoany kolonialne, które nie wykazują przemiany pokoleń , ale zamiast tego rozmnażają się bezpłciowo w procesie pączkowania. Zooidy to wielokomórkowe jednostki budujące kolonie. Pojedynczy pączek zwany pro-pączkiem inicjuje wzrost kolonii poprzez rozszczepienie. Każdy zooid jest produkowany tak, aby był genetycznie identyczny; jednak mutacje mogą zmienić ich funkcje i zwiększyć różnorodność zooidów w kolonii. Syfonofory są wyjątkowe, ponieważ pro-bud inicjuje produkcję różnych zooidów o określonych funkcjach. Funkcje i organizacja zooidów w koloniach znacznie się różnią między różnymi gatunkami; jednak większość kolonii jest ułożona obustronnie, grzbietową i brzuszną stroną łodygi. Łodyga to pionowa gałąź pośrodku kolonii, do której przyczepiają się zooidy. Zooidy zazwyczaj mają specjalne funkcje, a tym samym przyjmują określone wzory przestrzenne wzdłuż łodygi.
Morfologia ogólna
Syfonofory zazwyczaj wykazują jeden z trzech standardowych planów ciała. Plany ciała noszą nazwy Cystonecta, Physonecta i Calycophorae. Cystonekty mają długą łodygę z dołączonymi zooidami. Każda grupa zooidów ma gastrozooida. Gastrozooid ma mackę używaną do chwytania i trawienia pokarmu. Grupy mają również gonofory, które są wyspecjalizowane w rozmnażaniu. Używają pneumatoforu, pływaka wypełnionego gazem, na swoim przednim końcu i dryfują głównie na powierzchni wody. Physonects mają pneumatofor i nektosom, w którym znajdują się nektofory używane do napędu odrzutowego. Nektofory pompują wodę z powrotem, aby poruszać się do przodu. Calycophorans różnią się od cystonektów i fisonektów tym, że mają dwa nektofory i nie mają pneumatoforów.
Od czasu ich powstania w syfonoforach obserwuje się wzrost liczby typów zooidów. Naukowcy ustalili dwie możliwe hipotezy ewolucyjne dla tej obserwacji: 1. W miarę upływu czasu liczba typów zooidów wzrosła. 2. Ostatni wspólny przodek miał wiele typów zooidów, a różnorodność obserwowana dzisiaj wynika z utraty typów zooidów. Badania nie wykazują żadnych dowodów potwierdzających pierwszą hipotezę i pojawiły się dowody na poparcie drugiej.
- Zooidy
- Syfonofory mogą mieć zooidy, które są polipami lub meduzami. Jednak zooidy są wyjątkowe i mogą rozwijać się, aby pełnić różne funkcje.
- Nektofory
- Nektofory to meduzy, które pomagają w napędzaniu i poruszaniu się niektórych syfonoforów w wodzie. Są charakterystyczne dla physonectae i calycophores. Nektofory tych organizmów znajdują się w nektosomie, gdzie mogą koordynować pływanie kolonii. Zaobserwowano również, że nektofory działają w połączeniu ze strukturami rozrodczymi w celu zapewnienia napędu podczas odłączania się kolonii.
- przylistki
- Przylistki to zooidy, które są unikalne dla rzędu siphonophorae. Pełnią funkcję ochronną i utrzymują neutralną pływalność. Jednak przylistki nie występują we wszystkich gatunkach syfonoforów.
- Gastrozooidy
- Gastrozooidy to polipy, które rozwinęły funkcję wspomagającą odżywianie syfonoforów.
- Palpony
- Palpony to zmodyfikowane gastrozooidy, które działają w trawieniu poprzez regulację krążenia płynów żołądkowo-naczyniowych.
- Gonofory Gonofory
- to zooidy zaangażowane w procesy reprodukcyjne syfonoforów.
- Pneumatofory
- Obecność pneumatoforów charakteryzuje podgrupy Cystonectae i Physonectae. Są to wypełnione gazem pływaki, które znajdują się na przednim końcu kolonii tych gatunków. Działają, aby pomóc koloniom utrzymać orientację w wodzie. W podgrupie cystonectae pneumatofory pełnią dodatkową funkcję wspomagania flotacji organizmów. Syfonofory wykazujące tę cechę rozwijają strukturę we wczesnym rozwoju larwalnym poprzez wgłobienia spłaszczonej struktury planuli. Dalsze obserwacje gatunku siphonophor Nanomia bijuga wskazują, że cecha pneumatoforu potencjalnie działa również w celu wyczuwania zmian ciśnienia i regulacji chemotaksji u niektórych gatunków.
Dystrybucja i siedlisko
Obecnie Światowy Rejestr Gatunków Morskich (WoRMS) identyfikuje 175 gatunków syfonoforów. Mogą znacznie różnić się rozmiarem i kształtem, co w dużej mierze odzwierciedla środowisko, w którym żyją. Syfonofory są najczęściej organizmami pelagicznymi, jednak gatunki poziome są bentosowe . Mniejsze syfonofory ciepłowodne zwykle żyją w epipelagicznej i wykorzystują swoje macki do chwytania zooplanktonu i widłonogów . Większe syfonofory żyją w głębszych wodach, ponieważ są na ogół dłuższe i bardziej kruche i muszą unikać silnych prądów. Żywią się głównie większą zdobyczą. Większość syfonoforów żyje w głębinach morskich i można je znaleźć we wszystkich oceanach. Gatunki syfonoforowe rzadko zasiedlają tylko jedno miejsce. Jednak niektóre gatunki mogą być ograniczone do określonego zakresu głębokości i / lub obszaru oceanu.
Zachowanie
Ruch
Syfonofory wykorzystują metodę poruszania się podobną do napędu odrzutowego. Syfonofor to złożona kolonia zbiorcza składająca się z wielu nektoforów , które są klonalnymi osobnikami, które tworzą się przez pączkowanie i są genetycznie identyczne. W zależności od tego, gdzie każdy pojedynczy nektofor jest umieszczony w syfonoforze, ich funkcja jest różna. Ruch kolonialny jest determinowany przez poszczególne nektofory wszystkich stadiów rozwojowych. Mniejsze osobniki skupiają się w górnej części syfonoforu, a ich funkcją jest obracanie i dostosowywanie orientacji kolonii. Osoby stają się większe, im są starsze. Większe osobniki znajdują się u podstawy kolonii, a ich główną funkcją jest napęd ciągu. Te większe osobniki są ważne dla osiągnięcia maksymalnej prędkości kolonii. Każda jednostka jest kluczem do ruchu zbiorczej kolonii, a zrozumienie ich organizacji może pozwolić nam na poczynienie postępów w naszych własnych pojazdach z napędem wieloodrzutowym. Kolonialna organizacja syfonoforów, zwłaszcza w Nanomia bijuga zapewnia korzyści ewolucyjne. Duża liczba skoncentrowanych osobników pozwala na redundancję. Oznacza to, że nawet jeśli niektóre pojedyncze nektofory zostaną upośledzone funkcjonalnie, ich rola jest pomijana, więc kolonia jako całość nie jest negatywnie narażona. Welum, cienki pas tkanki otaczający otwór strumienia, również odgrywa rolę we wzorcach pływackich, co zostało wykazane w badaniach przeprowadzonych na wcześniej wspomnianym gatunku N. bijuga. Welum staje się mniejsze i bardziej okrągłe w czasie napędu do przodu w porównaniu z dużym welumem, które jest widoczne podczas okresów uzupełniania. Dodatkowo pozycja podniebienia zmienia się wraz z zachowaniami pływackimi; podniebienie wewnętrzne jest zakrzywione w dół w czasie natryskiwania, ale podczas uzupełniania podniebienie jest cofane z powrotem do nektoforu. Syfonofor Namonia bijuga również praktykuje migrację pionową diel , ponieważ pozostaje w głębinach morskich w ciągu dnia, ale wznosi się w nocy.
Drapieżnictwo i żerowanie
Syfonofory to drapieżne drapieżniki . Ich dieta składa się z różnych widłonogów, małych skorupiaków i małych ryb. Ogólnie rzecz biorąc, dieta silnie pływających syfonoforów składa się z mniejszej ofiary, a dieta słabo pływających syfonoforów składa się z większej ofiary. Większość syfonoforów ma gastrozooidy , które mają charakterystyczną mackę przymocowaną do podstawy zooida. Ta cecha strukturalna pomaga organizmom w łapaniu zdobyczy. Gatunki z dużymi gastrozooidami są w stanie zjadać ofiarę w szerokim zakresie rozmiarów. Podobny do wielu innych organizmów z gromady Cnidaria , wiele gatunków syfonoforów wykazuje parzące kapsułki nematocyst na gałęziach swoich macek zwanych tentilla. Nicienie są ułożone w gęste baterie po bokach namiotu. Kiedy syfonofor napotyka potencjalną ofiarę, jego tentillum reaguje na miejsce, w którym macki o długości 30–50 cm (12–20 cali) tworzą sieć, zmieniając swój kształt wokół ofiary. Następnie nematocysty wystrzeliwują miliony paraliżujących, a czasem śmiertelnych toksyn molekuły w uwięzionej ofierze, która jest następnie przenoszona do odpowiedniego miejsca w celu strawienia. Niektóre gatunki syfonoforów używają agresywnej mimikry za pomocą światła bioluminescencyjnego, więc ofiara nie może właściwie zidentyfikować drapieżnika.
Istnieją cztery rodzaje nematocyst w siphonophor tentilla: heteronemy, haplonemy, desmonemy i rhopalonemy. Heteronemy są największymi nematocystami i są kolcami na trzonie w pobliżu kanalików przymocowanych do środka syfonoforu. Haplonemy mają otwarte kanaliki z kolcami, ale nie mają wyraźnego trzonu. Jest to najczęstsza nematocysta wśród syfonoforów. Desmonemy nie mają kolców, ale zamiast tego kanaliki mają właściwości adhezyjne, które utrzymują zdobycz. Rhopalonemy to nematocysty z szerokimi kanalikami na zdobycz.
Ze względu na brak pożywienia w środowisku głębinowym większość gatunków syfonoforów funkcjonuje w taktyce „siedź i czekaj” na pożywienie. Galaretowaty plan ciała pozwala na elastyczność podczas łapania zdobyczy, ale galaretowate adaptacje są oparte na środowisku. Pływają w kółko, czekając, aż ich długie macki napotkają zdobycz. Ponadto syfonofory w grupie oznaczonej jako Erenna mają zdolność generowania bioluminescencji i czerwonej fluorescencji, podczas gdy jego macki drgają w sposób naśladujący ruchy małych skorupiaków i widłonogów. Działania te zachęcają ofiarę do zbliżenia się do syfonoforu, pozwalając jej złapać ją w pułapkę i strawić.
Reprodukcja
Sposoby rozmnażania sifonoforów różnią się między różnymi gatunkami i do dziś kilka trybów pozostaje nieznanych. Ogólnie rzecz biorąc, pojedyncza zygota rozpoczyna tworzenie kolonii zooidów. Zapłodnione jajo dojrzewa do pierwotniaka, który inicjuje proces pączkowania i tworzenia nowego zooida. Proces ten powtarza się, aż wokół centralnej łodygi utworzy się kolonia zooidów. W przeciwieństwie do tego, kilka gatunków rozmnaża się za pomocą polipów . Polipy mogą zawierać komórki jajowe i/lub plemniki i mogą być uwalniane do wody z tylnego końca syfonoforu. Polipy mogą następnie zostać zapłodnione poza organizmem.
Syfonofory wykorzystują gonofory do tworzenia gamet reprodukcyjnych . Gonofory są męskie lub żeńskie; jednak typy gonoforów w kolonii mogą się różnić w zależności od gatunku. Gatunki są charakteryzowane jako jednopienne lub dwupienne na podstawie ich gonoforów. jednopienne zawierają samce i samice gonoforów w jednej kolonii zooidów, podczas gdy gatunki dwupienne mają gonofory samców i samic osobno w różnych koloniach zooidów.
Bioluminescencja

Prawie wszystkie syfonofory mają zdolności bioluminescencyjne. Ponieważ organizmy te są niezwykle delikatne, rzadko obserwuje się je żywe. Uważa się, że bioluminescencja w syfonoforach wyewoluowała jako mechanizm obronny. , że syfonofory głębinowego rodzaju Erenna (znajdujące się na głębokościach między 1600 a 2300 metrów) wykorzystują swoje zdolności bioluminescencyjne również do ataku, jako przynęty do przyciągania ryb. Ten rodzaj jest jednym z nielicznych, który poluje raczej na ryby niż na skorupiaki. Organy bioluminescencyjne, zwane tentilla , u tych niewidomych osobników emitują czerwoną fluorescencję wraz z rytmicznym migoczącym wzorem, który przyciąga ofiarę, ponieważ przypomina mniejsze organizmy, takie jak zooplankton i widłonogi . Stwierdzono zatem, że wykorzystują luminescencję jako przynętę do przyciągania zdobyczy. Niektóre badania wskazują, że organizmy głębinowe nie są w stanie wykryć długich fal, a światło czerwone ma długą długość fali 680 nm. Jeśli tak jest, to Erenna nie wabi ryb i musi być inne wytłumaczenie. Jednak głębina morska pozostaje w dużej mierze niezbadana, a wrażliwość na światło czerwone u ryb, takich jak Cyclothone i myctophidae głębinowe ryb nie należy wyrzucać.
Przynęty bioluminescencyjne znajdują się w wielu różnych gatunkach syfonoforów i są używane z różnych powodów. Gatunki takie jak Agalma okeni Athorybia rosacea , Athorybia lucida i , Lychnafalma utricularia wykorzystują swoje przynęty jako urządzenie naśladujące przyciąganie zdobyczy. Uważa się, że A. rosacea naśladuje larwy ryb, A. lucida naśladuje domy larw, a L. utricularia naśladuje hydromedusę. Gatunek Resomia ornicephala wykorzystuje swoją zieloną i niebieską fluorescencyjną tentilla do przyciągania kryla, pomagając mu w konkurowaniu z innymi organizmami polującymi na tę samą zdobycz. Syfonofory z rodzaju Erenna używa bioluminescencyjnych przynęt otoczonych czerwoną fluorescencją, aby zwabić zdobycz i prawdopodobnie naśladować rybę z rodzaju Cyclothone . Ich ofiara jest zwabiona przez unikalne zachowanie związane z mackami. W młodości macki organizmów z Erenna zawierają tylko tkankę bioluminescencyjną, ale w miarę starzenia się organizmu w tych tkankach obecny jest również czerwony materiał fluorescencyjny.
Taksonomia
.jpg)
Organizmy z rzędu Siphonophorae zostały sklasyfikowane w typ Cnidaria i klasa Hydrozoa. Pokrewieństwa filogenetyczne syfonoforów cieszyły się dużym zainteresowaniem ze względu na dużą zmienność organizacji ich kolonii polipów i meduz. Kiedyś uważano, że jest to bardzo odrębna grupa, larw i cechy morfologiczne doprowadziły naukowców do przekonania, że syfonofory wyewoluowały z prostszych hydrozoanów kolonialnych, podobnych do tych z rzędów Anthoathecata i Leptothecata . W rezultacie są teraz zjednoczeni z tymi w podklasa Hydroidolina .
Wczesna analiza podzieliła syfonofory na 3 główne podgrupy w oparciu o obecność lub brak 2 różnych cech: pływające dzwony (nektofory) i pływaki (pneumatofory). Podgrupy składały się z Cystonectae, Physonectae i Calycorphores. Cystonectae miały pneumatofory, physonectae miały nektofory, a calycophores miały oba.
Eukariotyczny gen małej podjednostki jądrowej rybosomu 18S, eukariotyczny gen dużej podjednostki mitochondrialnej rybosomu 16S i analizy transkryptomu dodatkowo wspierają filogenetyczny podział Siphonophorae na 2 główne klady: Cystonectae i Codonophora. Podrzędy w Codonophora obejmują Physonectae (składające się z kladów Calycophorae i Euphysonectae), Pyrostephidae i Apolemiidae.
- Podrząd Calycophorae
- Abylidae Agassiz, 1862
- Clausophyidae Totton, 1965
- Diphyidae Quoy i Gaimard, 1827
- Hippopodiidae Kölliker, 1853
- Prayidae Kölliker, 1853
- Sphaeronectidae Huxley, 1859
- Tottonophyidae Pugh, Dunn i plamiak, 2018
- Podrząd Cystonectae
- Physaliidae Brandt, 1835
- Rhizophysidae Brandt, 1835
- Podrząd Physonectae
- Agalmatidae Brandt, 1834
- Apolemiidae Huxley, 1859
- Cordagalmatidae Pugh, 2016
- Erennidae Pugh, 2001
- Forskaliidae Haeckel, 1888
- Physophoridae Eschscholtz, 1829
- Pyrostephidae Moser, 1925
- Resomiidae Pugh, 2006
- Rhodaliidae Haeckel, 1888
- Stephanomiidae Huxley, 1859
Historia
Odkrycie
Carl Linneusz opisał pierwszego syfonoforu, portugalskiego żołnierza wojny , w 1758 roku. Tempo odkrywania gatunków syfonoforów było powolne w XVIII wieku, ponieważ znaleziono tylko cztery dodatkowe gatunki. W XIX wieku zaobserwowano 56 nowych gatunków dzięki wyprawom badawczym prowadzonym przez mocarstwa europejskie. Większość nowych gatunków znalezionych w tym okresie została zebrana w przybrzeżnych wodach powierzchniowych. Podczas wyprawy HMS Challenger zebrano różne gatunki syfonoforów. Ernsta Haeckla podjął próbę spisania wszystkich gatunków syfonoforów zebranych podczas tej wyprawy. Wprowadził 46 „nowych gatunków”; jednak jego praca była mocno krytykowana, ponieważ ostatecznie stwierdzono, że niektóre gatunki, które zidentyfikował, nie były syfonoforami. Niemniej jednak niektóre z jego opisów i rysunków (na zdjęciu poniżej) są uważane za przydatne przez współczesnych biologów. W XX wieku zaobserwowano tempo około 10 nowych odkryć gatunków na dekadę. Uważany za najważniejszego badacza syfonoforów, AK Totton wprowadził w połowie XX wieku 23 nowe gatunki syfonoforów.
6 kwietnia 2020 r. Schmidt Ocean Institute ogłosił odkrycie gigantycznego syfonoforu Apolemia w podwodnych kanionach w pobliżu wybrzeża Ningaloo , mierzącego 15 m (49 stóp) średnicy z pierścieniem o długości około 47 m (154 stóp), prawdopodobnie największego syfonoforu w historii nagrany.
Nie ma zapisów kopalnych syfonoforów, chociaż ewoluowały one i dostosowywały się przez długi czas. Ich gromada, Cnidaria , to starożytna linia, której początki sięgają ok. 640 milionów lat temu.
Syfonofory Haeckela
Ernst Haeckel opisał liczne syfonofory, a kilka tablic z jego Kunstformen der Natur (1904) przedstawia członków taksonu :

Płyta 7

Płyta 37
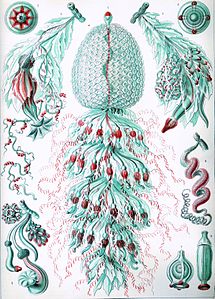
Płyta 59

Płyta 77




Dalsza lektura
- Mapstone, Gillian M. (2009). Siphonophora (Cnidaria, Hydrozoa) kanadyjskich wód Pacyfiku . Ottawa: NRC Research Press. ISBN 978-0-660-19843-9 .
- PinkTentacle.com (2008): Siphonophor: Superorganizm głębinowy (wideo) . Źródło 2009-MAJ-23.
Linki zewnętrzne
- Dunn, Casey (nd). „Syfonofory” . Bieżąca biologia . nie dotyczy 19 (6): R233-4. doi : 10.1016/j.cub.2009.02.009 . PMID 19321136 . Źródło 19 września 2014 r .
- Scubamedia.de (30 sierpnia 2013). „Tauchen w Norwegii - Kvasefjord” . YouTube . scubamedia.de. Zarchiwizowane od oryginału w dniu 2021-12-21 . Źródło 19 września 2014 r .
- Pinktentacle3 (22 grudnia 2008). „Syfonofor” . YouTube . Zarchiwizowane od oryginału w dniu 2021-12-21 . Źródło 19 września 2014 r .
- „Oszałamiająca obserwacja syfonoforu” . Nautilus Live: Eksploruj ocean NA ŻYWO z dr Robertem Ballardem i Korpusem Eksploracyjnym . Fundusz Eksploracji Oceanów. 27 czerwca 2014 . Źródło 18 września 2014 r .
- „Syfonofor głębinowy” (10 kwietnia 2017) YouTube. Sfotografowany przez NOAA Okeanos Explorer 14 marca 2017 r. Na wysokości 1560 metrów na zachód od kompleksu Winslow Reef. Źródło 28 stycznia 2018 r.