Morfologia sinic

Morfologia sinic odnosi się do formy lub kształtu sinic. Cyjanobakterie to duża i zróżnicowana gromada bakterii charakteryzująca się unikalną kombinacją barwników i zdolnością do fotosyntezy tlenowej .
Cyjanobakterie często żyją w skupiskach kolonialnych , które mogą przybierać różne formy. Wśród wielu gatunków cyjanobakterii szczególnie interesujące są te, które żyją kolonialnie w wydłużonych strukturach przypominających włosy, znanych jako trichomy . Te nitkowate gatunki mogą zawierać setki do tysięcy komórek. Często dominują w górnych warstwach mat mikrobiologicznych występujących w ekstremalnych środowiskach, takich jak gorące źródła , woda hipersoli , pustynie i regiony polarne, a także są szeroko rozpowszechnione w bardziej przyziemnych środowiskach.
Wiele gatunków nitkowatych jest również ruchliwych , ślizgając się wzdłuż swojej długiej osi i wykazując fotoruch , za pomocą którego trichom moduluje swój ślizg zgodnie z padającym światłem. Stwierdzono, że to ostatnie odgrywa ważną rolę w kierowaniu trichomów do optymalnych warunków oświetleniowych, które mogą albo zahamować komórki, jeśli padające światło jest zbyt słabe, albo uszkodzić komórki, jeśli jest zbyt mocne.
Różnorodność form
Cyjanobakterie wykazują niezwykłą zmienność pod względem morfologii: od jednokomórkowych i kolonialnych do wielokomórkowych form nitkowatych. Wielkość ich komórek waha się od mniej niż 1 µm średnicy (picocyjanobakterie) do 100 µm (niektóre formy tropikalne z rodzaju Oscillatoria )
Formy nitkowate wykazują funkcjonalne różnicowanie komórek, takie jak heterocysty (do wiązania azotu), akinety (komórki w stanie spoczynku) i hormogonia (rozrodcze, ruchliwe włókna). Te, wraz z posiadanymi przez nie połączeniami międzykomórkowymi, są uważane za pierwsze oznaki wielokomórkowości.
Wiele cyjanobakterii tworzy ruchome włókna komórek, zwane hormogoniami , które oddalają się od głównej biomasy, by pączkować i tworzyć nowe kolonie gdzie indziej. Komórki w hormogonium są często cieńsze niż w stanie wegetatywnym, a komórki na obu końcach ruchliwego łańcucha mogą być zwężone. Aby oderwać się od kolonii macierzystej, hormogonium często musi rozerwać słabszą komórkę we włóknie, zwaną nekrydium.

Jednokomórkowe i kolonialne cyjanobakterie słupki skali około 10 µm



Cyjanobakterie związane z tufami Schizothrix calcicola , Gloeocapsa , Coccochloris , Microcoleus vaginatus i Rivularia
Kolonialne i jednokomórkowe
W siedliskach wodnych sinice jednokomórkowe są uważane za ważną grupę pod względem liczebności, różnorodności i charakteru ekologicznego. Jednokomórkowe cyjanobakterie mają kuliste, jajowate lub cylindryczne komórki, które mogą agregować w nieregularne lub regularne kolonie związane razem przez macierz śluzową ( śluz ) wydzielaną podczas wzrostu kolonii. W zależności od gatunku liczba komórek w każdej kolonii może wahać się od dwóch do kilku tysięcy.
Każda pojedyncza komórka (każda pojedyncza sinica) ma zazwyczaj grubą, galaretowatą ścianę komórkową . Brakuje im wici , ale hormogonia niektórych gatunków może poruszać się, ślizgając się po powierzchniach.
![Merismopedia forms rectangular colonies held together by a mucilaginous matrix. Species in this genus divide in only two directions, creating a characteristic grid-like pattern arranged in rows and flats.[19]](//upload.wikimedia.org/wikipedia/commons/thumb/e/e6/Merismopedia.jpg/393px-Merismopedia.jpg)
Merismopedia tworzy prostokątne kolonie utrzymywane razem przez śluzowatą matrycę . Gatunki z tego rodzaju dzielą się tylko w dwóch kierunkach, tworząc charakterystyczny wzór przypominający siatkę, ułożony w rzędy i mieszkania.
![Merismopedia forms rectangular colonies held together by a mucilaginous matrix. Species in this genus divide in only two directions, creating a characteristic grid-like pattern arranged in rows and flats.[19]](https://commons.wikimedia.org/wiki/File:Merismopedia.jpg)

Kolonie Nostoc pruniforme „galaretki”

Sinice kolonialne Stratonostoc
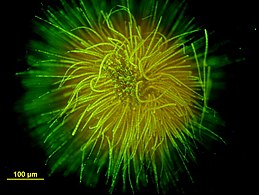
Kulista kolonia Gloeotrichia echinulata

Kolonia cyjanobakterii Lyngbya majuscula




Nitkowate i wielokomórkowe
![Example of filamentous cyanobacteria structure (Oscillatoria lutea) showing a reticulate pattern [20]](//upload.wikimedia.org/wikipedia/commons/thumb/b/b9/Filamentous_cyanobacteria_structure_of_Oscillatoria_lutea.jpg/359px-Filamentous_cyanobacteria_structure_of_Oscillatoria_lutea.jpg)
Przykład struktury nitkowatych cyjanobakterii ( Oscillatoria lutea ) przedstawiający wzór siatkowaty
![Example of filamentous cyanobacteria structure (Oscillatoria lutea) showing a reticulate pattern [20]](https://commons.wikimedia.org/wiki/File:Filamentous_cyanobacteria_structure_of_Oscillatoria_lutea.jpg)
Niektóre gatunki nitkowate mogą różnicować się w kilka różnych typów komórek :
- komórki wegetatywne – normalne komórki fotosyntetyzujące, które powstają w sprzyjających warunkach wzrostu
- akinetes – odporne na klimat zarodniki, które mogą tworzyć się, gdy warunki środowiskowe stają się trudne
- grubościenne heterocysty – które zawierają enzym azotazę niezbędną do wiązania azotu w środowisku beztlenowym ze względu na swoją wrażliwość na tlen.
Wiele wielokomórkowych nitkowatych form Oscillatoria jest zdolnych do ruchu falowego; włókno oscyluje tam iz powrotem. W słupach wody niektóre cyjanobakterie unoszą się, tworząc pęcherzyki gazu , jak u archeonów . Te pęcherzyki nie są organellami jako takimi. Nie są otoczone błonami lipidowymi , lecz otoczką białkową.

Mikrofotografie cyjanobakterii nitkowatych tworzących wiązki A–C: Microcoleus steenstrupii D–E: Tolypothrix desertorum F: Scytonema cf. calcicola G: S. por. calcicola H: S. por. c alcicola Pasek skali =10 µm
![Anabaena is used as a model organism to study simple vision[25]](//upload.wikimedia.org/wikipedia/commons/thumb/d/d1/Necklace_of_Mermaid.tif/lossless-page1-260px-Necklace_of_Mermaid.tif.png)
Anabaena jest używana jako organizm modelowy do badania prostego widzenia

Spiralne włókna cyjanobakterii
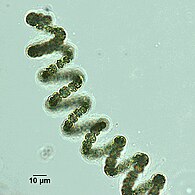
Filament spiralny z Dolichospermum

Lyngbya tworzą długie, nierozgałęzione włókna wewnątrz sztywnych śluzowatych osłonek, które mogą tworzyć sploty lub maty, zmieszane z innymi gatunkami fitoplanktonu

![Anabaena is used as a model organism to study simple vision[25]](http://upload.wikimedia.org/wikipedia/commons/thumb/d/d1/Necklace_of_Mermaid.tif/lossless-page1-260px-Necklace_of_Mermaid.tif.png)



Rozgałęziony
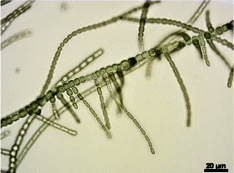
Prawdziwy rozgałęziony fenotyp kolonii Fischerella thermalis


Heterocysty
Heterocysty to wyspecjalizowane komórki wiążące azot, utworzone podczas głodu azotu przez niektóre nitkowate cyjanobakterie, takie jak Nostoc punctiforme , Cylindrospermum stagnale i Anabaena sphaerica . Wiążą azot z atmosferycznego N 2 za pomocą enzymu azotazy , aby dostarczyć komórkom we włóknie azot do biosyntezy.
Ruch

Cyjanobakterie są wszechobecne, znajdując siedliska w większości zbiorników wodnych iw ekstremalnych środowiskach, takich jak regiony polarne, pustynie, jeziora solankowe i gorące źródła. Wyewoluowały również zaskakująco złożone zachowania zbiorowe, które leżą na granicy między życiem jednokomórkowym i wielokomórkowym. Na przykład cyjanobakterie nitkowate żyją w długich łańcuchach komórek, które łączą się w większe struktury, w tym biofilmy , biomaty i stromatolity . Te duże kolonie zapewniają sztywne, stabilne i długotrwałe środowisko dla swoich społeczności bakterii. Ponadto biofilmy na bazie sinic mogą być wykorzystywane jako bioreaktory do produkcji szerokiej gamy chemikaliów, w tym biopaliw, takich jak biodiesel i etanol. Jednak pomimo ich znaczenia dla historii życia na Ziemi oraz ich potencjału komercyjnego i środowiskowego, pozostają podstawowe pytania dotyczące tego, jak nitkowate cyjanobakterie poruszają się, reagują na swoje środowisko i samoorganizują się w kolektywne wzorce i struktury.
Wszystkie znane cyjanobakterie nie mają wici ; jednak wiele gatunków nitkowatych porusza się po powierzchniach ślizgając się, formą poruszania się, w której nie widać żadnych fizycznych wyrostków wspomagających ruch. Rzeczywisty mechanizm szybowania nie jest w pełni poznany, chociaż od jego odkrycia upłynęło ponad sto lat. Jedna z teorii sugeruje, że w ruchu ślizgowym cyjanobakterii pośredniczy ciągłe wydzielanie polisacharydów przez pory w poszczególnych komórkach. Inna teoria sugeruje, że ruch ślizgowy obejmuje użycie pilusów typu IV , polimerowych zespołów pilinu białkowego , jako silników napędowych ruchu. Jednak nie jest jasne, w jaki sposób działanie tych pilusów prowadziłoby do ruchu, przy czym niektórzy sugerują, że wycofują się, podczas gdy inni sugerują, że pchają, aby generować siły. Inni uczeni zasugerowali, że fale powierzchniowe generowane przez kurczenie się warstwy włóknistej są mechanizmem ruchu ślizgowego w oscylatoriach . Niedawne prace sugerują również, że fluktuacje kształtu i siły kapilarne mogą być zaangażowane w ruch ślizgowy.
Poprzez zbiorową interakcję cyjanobakterie nitkowate samoorganizują się w kolonie lub biofilmy, społeczności symbiotyczne występujące w wielu różnych niszach ekologicznych. Ich struktury zbiorowe na większą skalę charakteryzują się różnorodnymi kształtami, w tym wiązkami, wirami i wzorami siatkowymi . Podobne wzorce zaobserwowano w zapisach kopalnych. Wiadomo, że w przypadku cyjanobakterii nitkowatych mechanika włókien przyczynia się do samoorganizacji, na przykład w określaniu, w jaki sposób jedno włókno ugnie się w kontakcie z innymi włóknami lub przeszkodami. Ponadto biofilmy i biomaty wykazują pewne wyjątkowo zachowane właściwości makromechaniczne, zwykle zachowując się jak materiały lepkosprężyste z czasem relaksacji około 20 minut.
Cyjanobakterie mają ścisłe wymagania dotyczące światła. Zbyt mało światła może skutkować niewystarczającą produkcją energii, a u niektórych gatunków może spowodować, że komórki będą uciekać się do oddychania heterotroficznego. Zbyt dużo światła może hamować komórki, zmniejszać wydajność fotosyntezy i powodować uszkodzenia poprzez blaknięcie. Promieniowanie UV jest szczególnie zabójcze dla cyjanobakterii, a normalny poziom nasłonecznienia jest w niektórych przypadkach znacznie szkodliwy dla tych mikroorganizmów.
Cyjanobakterie nitkowate, które żyją w matach mikrobiologicznych, często migrują pionowo i poziomo w macie, aby znaleźć optymalną niszę, która równoważy ich zapotrzebowanie na światło do fotosyntezy z wrażliwością na fotouszkodzenia. Na przykład nitkowate cyjanobakterie Oscillatoria sp. i Spirulina subsalsa znalezione w hipersalinowych matach bentosowych Guerrero Negro w Meksyku migrują w dół do niższych warstw w ciągu dnia, aby uciec przed intensywnym światłem słonecznym, a następnie wznoszą się na powierzchnię o zmierzchu. W przeciwieństwie do tego populacja Microcoleus chthonoplastes znaleziona w matach hipersalinowych w Salin-de-Giraud , Camargue, Francja migruje do górnej warstwy maty w ciągu dnia i jest równomiernie rozprowadzana przez matę w nocy. Eksperyment in vitro z użyciem P. uncinatum również wykazał tendencję tego gatunku do migracji w celu uniknięcia szkodliwego promieniowania. Te migracje są zwykle wynikiem pewnego rodzaju ruchu fotograficznego, chociaż inne formy taksówek również mogą odgrywać pewną rolę.
Wiele gatunków cyjanobakterii jest zdolnych do szybowania. Szybowanie jest formą ruchu komórki, która różni się od pełzania lub pływania tym, że nie opiera się na żadnym oczywistym narządzie zewnętrznym ani zmianie kształtu komórki i występuje tylko w obecności podłoża . Szybowanie w cyjanobakterii nitkowatej wydaje się być napędzane mechanizmem „strumienia śluzu”, w którym komórki wytłaczają żel, który szybko się rozszerza, gdy się nawadnia, zapewniając siłę napędową, chociaż niektóre jednokomórkowe cyjanobakterie używają pilusów typu IV do szybowania . Poszczególne komórki włoska mają dwa zestawy porów do wytłaczania śluzu. Każdy zestaw jest zorganizowany w pierścień w przegrodach komórkowych i wytłacza śluz pod ostrym kątem. Zestawy wytłaczają szlam w przeciwnych kierunkach, więc tylko jeden zestaw prawdopodobnie zostanie aktywowany podczas szybowania. Alternatywna hipoteza głosi, że komórki wykorzystują elementy kurczliwe, które wytwarzają falowanie przebiegające po powierzchni wewnątrz rurki śluzowej jak dżdżownica. Trichomy obracają się w sposób spiralny, którego kąt odpowiada kątowi nachylenia kurczliwych trichomów Castenholza.


Wydaje się, że komórki koordynują swój kierunek poślizgu za pomocą potencjału elektrycznego, który ustala polaryzację we włoskach, a tym samym ustanawia „głowę” i „ogon”. Trichomy zwykle losowo odwracają swoją polaryzację ze średnim okresem rzędu minut do godzin. Wiele gatunków tworzy również półsztywną osłonę, która pozostaje w postaci pustej rurki, gdy trichom porusza się do przodu. Kiedy trichom zmienia kierunek, może wrócić do pochwy lub wyrwać się.
Oscillatoria to rodzaj nitkowatych cyjanobakterii, których nazwa pochodzi od oscylacji w ich ruchu. Włókna w koloniach przesuwają się tam iz powrotem względem siebie, aż cała masa zostanie przestawiona na źródło światła. Oscillatoria jest głównie niebiesko-zielona lub brązowo-zielona i jest powszechnie spotykana w poidłach. Rozmnaża się przez fragmentację , tworząc długie włókna komórek, które mogą pękać na fragmenty zwane hormogoniami . Hormogonia może następnie wyrosnąć na nowe, dłuższe włókna.
Eksperyment z cyjanografem Hädera

W 1987 roku Häder wykazał, że trichomy mogą dość precyzyjnie pozycjonować się w swoim środowisku dzięki fotoruchowi . W eksperymencie cyjanograficznym Hädera negatyw fotograficzny jest rzutowany na szalkę Petriego zawierającą kulturę Phormidium uncinatum . Po kilku godzinach trichomy przesuwają się z ciemniejszych obszarów na jaśniejsze, tworząc fotograficzny pozytyw kultury. Eksperyment pokazuje, że fotoruch jest skuteczny nie tylko w przypadku dyskretnych pułapek świetlnych, ale także w przypadku drobnoziarnistych, stale zróżnicowanych pól świetlnych.