Pilus

Pilus ( łac. „włosy”; liczba mnoga: pilus ) to podobny do włosów wyrostek występujący na powierzchni wielu bakterii i archeonów . Terminy pilus i fimbria (łac. „frędzle”; liczba mnoga: fimbriae ) mogą być używane zamiennie, chociaż niektórzy badacze rezerwują termin pilus dla wyrostka robaczkowego wymaganego do koniugacji bakteryjnej . Wszystkie pilusy koniugacyjne składają się głównie z pilinów – białek włóknistych , które są oligomerami .
Na powierzchni bakterii i archeonów mogą istnieć dziesiątki takich struktur. Niektóre bakterie, wirusy lub bakteriofagi przyczepiają się do receptorów na pilusach na początku ich cyklu reprodukcyjnego .
Pili są antygenowe . Są one również kruche i stale zastępowane, czasem pilusami o różnym składzie, co skutkuje zmienioną antygenowością. Specyficzne reakcje żywiciela na stare struktury pilusów nie są skuteczne w przypadku nowej struktury. Rekombinacja między genami niektórych (ale nie wszystkich) pilusów koduje regiony zmienne (V) i stałe (C) pilusów (podobnie jak immunoglobulin ). Jako główne determinanty antygenowe, czynniki wirulencji i czynniki bezkarności na powierzchni komórek wielu gatunków bakterii Gram-ujemnych i niektórych Gram-dodatnich, w tym Enterobacteriaceae , Pseudomonadaceae i Neisseriaceae , badanie pilusów jako organelli adhezyjnych i składników szczepionek wzbudziło duże zainteresowanie. Pierwsze szczegółowe badanie pilusów zostało przeprowadzone przez Brintona i współpracowników, którzy wykazali istnienie dwóch odrębnych faz w obrębie jednego szczepu bakteryjnego: włochatego (p+) i nie włochatego)
Typy według funkcji
Kilka nazw nadano różnym typom pilusów ze względu na ich funkcję. Klasyfikacja nie zawsze pokrywa się z typami strukturalnymi lub ewolucyjnymi, ponieważ zachodzi ewolucja zbieżna .
Pili koniugacyjne
Pilusy koniugacyjne umożliwiają przenoszenie DNA między bakteriami w procesie koniugacji bakteryjnej . Nazywa się je czasami „pilusami płciowymi”, analogicznie do rozmnażania płciowego , ponieważ umożliwiają wymianę genów poprzez tworzenie „par godowych”. Być może najlepiej zbadanym jest pilus F Escherichia coli , zakodowany przez czynnik płciowy F.
Pilus płciowy ma zwykle średnicę od 6 do 7 nm . Podczas koniugacji pilus wyłaniający się z bakterii dawcy chwyta bakterię biorcę w sidła, przyciąga ją do siebie i ostatecznie wyzwala tworzenie mostka godowego , który ustanawia bezpośredni kontakt i tworzenie kontrolowanych porów, które umożliwiają przeniesienie DNA od dawcy do odbiorca. Zazwyczaj przenoszone DNA składa się z genów wymaganych do wytworzenia i przeniesienia pilusów (często zakodowanych na plazmidzie ) , podobnie jak rodzaj samolubnego DNA ; jednak inne fragmenty DNA są często przenoszone razem, co może skutkować rozpowszechnieniem cech genetycznych w całej populacji bakterii, takich jak oporność na antybiotyki . Nie wszystkie bakterie mogą tworzyć pilusy koniugacyjne, ale koniugacja może wystąpić między bakteriami różnych gatunków. [ potrzebne źródło ]

Hipertermofilne archeony kodują pilusy strukturalnie podobne do bakteryjnych pilusów koniugacyjnych. Jednak w przeciwieństwie do bakterii, gdzie aparat koniugacyjny zazwyczaj pośredniczy w przenoszeniu ruchomych elementów genetycznych, takich jak plazmidy lub transpozony, maszyneria koniugacyjna hipertermofilnych archeonów, zwana Ced (Crenarchaeal system wymiany DNA) i Ted (Thermoproteales system wymiany DNA ), wydaje się być odpowiedzialny za przenoszenie komórkowego DNA między przedstawicielami tego samego gatunku. Sugerowano, że w tych archeonach mechanizm koniugacji został w pełni udomowiony w celu promowania naprawy DNA poprzez rekombinację homologiczną, a nie rozprzestrzenianie się ruchomych elementów genetycznych.
fimbrie

Fimbria ( z łaciny „frędzle”, liczba mnoga fimbriae ) to termin używany na określenie krótkiego pilusa, który służy do przyczepiania bakterii do powierzchni, czasami nazywanego również „pilusem mocującym”. Termin „fimbria” może odnosić się do wielu różnych (strukturalnych) typów pilusów, ponieważ wiele różnych typów pilusów było używanych do adhezji, co jest przypadkiem zbieżnej ewolucji . System Gene Ontology nie traktuje fimbrii jako odrębnego rodzaju przydatków, zamiast tego używa ogólnego typu pilus (GO: 0009289).
Ten wyrostek ma średnicę od 3 do 10 nanometrów i może mieć długość nawet kilku mikrometrów. Fimbrie są wykorzystywane przez bakterie do przylegania do siebie i przylegania do komórek zwierzęcych i niektórych przedmiotów nieożywionych. Bakteria może mieć nawet 1000 fimbrii. Fimbrie są widoczne tylko pod mikroskopem elektronowym . Mogą być proste lub elastyczne.
Fimbriae posiadają adhezyny , które przyczepiają je do pewnego rodzaju podłoża, dzięki czemu bakterie mogą wytrzymać siły ścinające i uzyskać składniki odżywcze. Na przykład E. coli wykorzystuje je do przyłączania się do receptorów mannozy .
Niektóre bakterie tlenowe tworzą bardzo cienką warstwę na powierzchni kultury bulionowej . Ta warstwa, zwana błonką , składa się z wielu bakterii tlenowych, które przylegają do powierzchni za pomocą fimbrii. W ten sposób fimbrie pozwalają bakteriom tlenowym przebywać zarówno na bulionie, z którego pobierają składniki odżywcze, jak iw pobliżu powietrza.
Fimbrie są wymagane do tworzenia biofilmu , ponieważ przyczepiają bakterie do powierzchni żywiciela w celu kolonizacji podczas infekcji. Fimbrie znajdują się albo na biegunach komórki, albo są równomiernie rozmieszczone na całej jej powierzchni.
Termin ten był również używany w luźnym sensie w odniesieniu do wszystkich pilusów przez tych, którzy używają słowa „pilus” w odniesieniu konkretnie do pilusów płciowych.
Typy według systemu montażu lub struktury
Przenosić
Rodzina Tra (transfer) obejmuje wszystkie znane pilusy płciowe (od 2010 r.). Są one związane z układem wydzielniczym typu IV (T4SS). Można je podzielić na typ F-podobny (po F-pilus) i typ P-podobny. Podobnie jak ich odpowiedniki wydzielnicze, pilus wstrzykuje materiał, w tym przypadku DNA, do innej komórki.
Pilusy typu IV

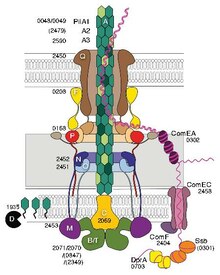
Niektóre pilusy, zwane pilusami typu IV (T4P), generują siły ruchowe . Zewnętrzne końce pilusów przylegają do stałego podłoża, albo do powierzchni, do której bakteria jest przyczepiona, albo do innych bakterii. Następnie, gdy pilusy kurczą się, ciągną bakterię do przodu jak hak. Ruch wytwarzany przez pilusy typu IV jest zwykle gwałtowny, dlatego nazywa się go ruchliwością drgawkową , w przeciwieństwie do innych form ruchliwości bakteryjnej, takich jak ta wytwarzana przez wici . Jednak niektóre bakterie, na przykład Myxococcus xanthus , wykazują ruchliwość ślizgową . Pilusy bakteryjne typu IV mają podobną budowę do białek składowych archaelli (wici archeonów) i oba są spokrewnione z systemem wydzielniczym typu II (T2SS); łączy je grupa filamentów typu IV . Oprócz archaelli wiele archeonów wytwarza klejące pilusy typu 4, które umożliwiają komórkom archeonów przyleganie do różnych podłoży. N-końcowe alfa-helikalne części pilin i archaelin typu 4 archeonów są homologiczne do odpowiednich regionów bakteryjnego T4P; jednakże wydaje się, że domeny bogate w C-końcowe nici beta nie są spokrewnione w pilinach bakteryjnych i archeologicznych.
Transformacja genetyczna to proces, w którym bakteryjna komórka biorcy pobiera DNA z sąsiedniej komórki i integruje ten DNA ze swoim genomem poprzez rekombinację homologiczną . W Neisseria meningitidis (zwanym także meningokokiem) transformacja DNA wymaga obecności krótkich sekwencji wychwytu DNA (DUS), które składają się z 9-10 monomerów rezydujących w regionach kodujących DNA dawcy. W specyficznym rozpoznaniu DUS pośredniczy pilin typu IV . Pilusy menningokokowe typu IV wiążą DNA przez mniejszy pilin ComP poprzez elektropozytywny pasek, który, jak przewiduje się, zostanie wyeksponowany na powierzchni włókna. ComP wykazuje wyjątkową preferencję wiązania dla selektywnych DUS. Rozmieszczenie DUS w N. meningitides faworyzuje określone geny, co sugeruje, że istnieje tendencja do genów zaangażowanych w konserwację i naprawę genomu.
Ta rodzina została pierwotnie zidentyfikowana jako „fimbrie typu IV” na podstawie ich wyglądu pod mikroskopem. Ta klasyfikacja przetrwała, ponieważ odpowiada kladowi.
Fimbrie typu 1
Inny typ to fimbrie typu 1. Zawierają adhezyny FimH na „końcówkach”. Szlak opiekuńczo-wprowadzający jest odpowiedzialny za przemieszczanie wielu typów fimbrii z komórki, w tym fimbrii typu 1 i fimbrii P.
Curli

„Bakterie Gram-ujemne gromadzą funkcjonalne włókna powierzchniowe amyloidu zwane curli ”. Curli to rodzaj fimbrii. Curli składają się z białek zwanych lokami. Niektóre zaangażowane geny to CsgA , CsgB , CsgC , CsgD , CsgE , CsgF i CsgG .
Zjadliwość
Pilusy są odpowiedzialne za zjadliwość patogennych szczepów wielu bakterii, w tym E. coli , Vibrio cholerae i wielu szczepów Streptococcus . Dzieje się tak, ponieważ obecność pilusów znacznie zwiększa zdolność bakterii do wiązania się z tkankami ciała, co z kolei zwiększa szybkość replikacji i zdolność do interakcji z organizmem gospodarza. Jeśli gatunek bakterii ma wiele szczepów, ale tylko niektóre są patogenne, prawdopodobne jest, że szczepy patogenne będą miały pilusy, podczas gdy szczepy niepatogenne nie.
Rozwój pilusów przywiązania może następnie skutkować rozwojem dalszych cech zjadliwości. Fimbrie są jednym z głównych mechanizmów wirulencji bakterii E. coli , Bordetella pertussis , Staphylococcus i Streptococcus . Ich obecność znacznie zwiększa zdolność bakterii do przyczepiania się do żywiciela i wywoływania choroby. Niepatogenne szczepy V. cholerae jako pierwsze wyewoluowały pilusy, co pozwoliło im wiązać się z ludzkimi tkankami i tworzyć mikrokolonie . Te pilusy następnie służyły jako miejsca wiązania dla lizogenny bakteriofag , który przenosi chorobotwórczą toksynę . Gen tej toksyny, po włączeniu do genomu bakterii, ulega ekspresji, gdy ulega ekspresji gen kodujący pilus (stąd nazwa „pilus za pośrednictwem toksyny”).
Zobacz też
Linki zewnętrzne
- Sex + Pilus w US National Library of Medicine Medical Subject Headings (MeSH)
- Bacterial + Pilus w US National Library of Medicine Medical Subject Headings (MeSH)
- Fimbriae+Proteins w US National Library of Medicine Medical Subject Headings (MeSH)
| Mikrobiologia medyczna |
|||||||
|---|---|---|---|---|---|---|---|
|
Biochemia i ekologia |
|
||||||
| Kształt | |||||||
| Struktura |
|
||||||
|
Taksonomia i ewolucja |
|||||||