Angomonas deanei
|
|
| Angomonas deanei | |
|---|---|
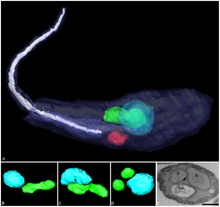
| Trójwymiarowa rekonstrukcja Angomonas deanei zawierająca endosymbiont bakteryjny (zielony) w pobliżu jądra (niebieski). | |
| Klasyfikacja naukowa | |
| Gromada: | Euglenozoa |
| Klasa: | kinetoplastyka |
| Zamówienie: | trypanosomatyda |
| Rodzaj: | Angomony |
| Gatunek: |
A. deanei
|
| Nazwa dwumianowa | |
|
Angomonas deanei (Carvalho, 1973) Teixeira i Camargo, 2011 |
|
| Synonimy | |
|
Crithidia deanei Carvalho, 1973 |
|
Angomonas deanei to wiciowy pierwotniak świdrowców . Jako pasożyt bezwzględny zaraża przewód pokarmowy owadów , a z kolei jest żywicielem bakterii symbiotycznych . Bakteryjny endosymbiont Ca. „ Kinetoplastibacterium crithidii ” utrzymuje stały , wzajemny związek z pierwotniakiem, tak że nie jest już w stanie samodzielnie się rozmnażać i przetrwać. Symbioza, odkryta następnie w różnym stopniu u innych protistów, takich jak Strigomonas culicis , Novymonas esmeraldas , Diplonema japonicum i Diplonema aggregatum , jest uważana za dobry model do zrozumienia ewolucji eukariontów z prokariotów oraz pochodzenia organelli komórkowych (tj. symbiogeneza ).
Gatunek został po raz pierwszy opisany jako Crithidia deanei w 1973 roku przez brazylijskiego parazytologa Aurorę LM Carvalho. Analiza filogenetyczna przeprowadzona w 2011 roku wykazała, że należy do rodzaju Angomonas , stając się tym samym Angomonas deanei . Symbiotyczna bakteria jest członkiem β-proteobakterii , która wywodzi się od wspólnego przodka z rodzaju Bordetella lub, co bardziej prawdopodobne, Taylorella . Te dwa organizmy polegały na sobie tak bardzo, że bakteria nie może się rozmnażać, a pierwotniak nie może już zarażać owadów, gdy są izolowane.
Odkrycie
Angomonas deanei został pierwotnie opisany jako Crithidia deanei . W 1973 roku brazylijska absolwentka Aurora Luiza de Moura Carvalho z Universidade Federal de Goiás odkryła ten gatunek na podstawie swoich badań nad pasożytami jelitowymi owadów- zabójców w Goiás. W następnym roku poinformował, że robak Zelus leucogrammus , którego odkrył, nie był naturalnie zainfekowany przez pierwotniaka, ale został nabyty od innych owadów. W tym samym czasie zespół badawczy z Universidade de Brasilia przedstawił właściwości biochemiczne i szczegóły strukturalne na podstawie transmisyjnej mikroskopii elektronowej. Odkryli, że zawiera on endosymbiont, opisując go jako „prawdopodobnie bakteryjnego”, który dostarcza „niezbędnych składników odżywczych trypanosomatydu”. Bakteryjny charakter endosymbiontu został potwierdzony w 1977 r., kiedy wykazano, że można go zabić przez leczenie antybiotykiem chloramfenikolem i że pomaga gospodarzowi w syntezie aminokwasu argininy z ornityny.
Gdy badano więcej szczegółów strukturalnych i molekularnych, rozróżnienie A. deanei od innych gatunków Crithidia stało się bardziej wyraźne. W 1991 roku Maria Auxiliadora de Sousa i Suzana Corte-Real z Instituto Oswaldo Cruz zaproponowały nowy rodzaj Angomonas dla gatunku. Badania filogenetyczne przeprowadzone przez Martę MG Teixeira i Erneya P. Camargo na Uniwersytecie w São Paulo wraz z ich współpracownikami w 2011 r. potwierdziły nową nazwę gatunku A. deanei wraz z opisem nowego spokrewnionego gatunku A. ambiguus , który również zawiera ten sam endosymbiont bakteryjny .
Struktura

Ciało Angomonas deanei ma kształt eliptyczny, z wydatną wicią przypominającą ogon na tylnym końcu ułatwiającą poruszanie się. Endosymbiont bakteryjny znajduje się wewnątrz jego ciała i jest otoczony przez dwie błony komórkowe typowe dla bakterii Gram-ujemnych , ale jego błona komórkowa ma niezwykłe cechy, takie jak obecność fosfatydylocholiny , głównego lipidu błonowego (nietypowego dla błon bakteryjnych) oraz wysoce zredukowana warstwa peptydoglikanu , która wykazuje zmniejszoną lub brak sztywnej ściany komórkowej . Błona komórkowa żywiciela pierwotniaka zawiera 18-domenową β-beczułkę porynową , która jest charakterystycznym białkiem bakterii Gram-ujemnych i nietypowym dla eukariontów. Ponadto zawiera kardiolipinę i fosfatydylocholinę jako główne fosfolipidy , podczas gdy sterole są nieobecne. Kardiolipina jest typowym lipidem błon bakteryjnych; z drugiej strony fosfatydylocholina występuje głównie w symbiotycznych prokariotach komórek eukariotycznych. W celu adaptacji symbiotycznej gospodarz pierwotniak przeszedł zmiany, takie jak zmniejszenie pręcika paraflagellarnego, który jest wymagany do pełnej ruchliwości wici bakteryjnej. Jednak gen pręcika paraflagellarnego PFR1 jest w pełni funkcjonalny. Brakuje również intronów i transkrypcji długich policistronowych mRNA wymaganych przez inne eukarionty do złożonej aktywności genów. Cały jego genom jest rozmieszczony w 29 chromosomach i zawiera 10 365 genów kodujących białka, 59 transferowych RNA, 26 rybosomalnych RNA i 62 niekodujące RNA.
Podczas gdy pierwotniak ma swoje oddzielne mitochondria, które zapewniają system transportu elektronów do produkcji energii komórkowej, cząsteczki ATP są wytwarzane przez jego glikosomy . Wiadomo, że bakteria dostarcza gospodarzowi niezbędnych składników odżywczych. Syntetyzuje aminokwasy, witaminy, zasady azotowe i hem dla pierwotniaków. Hem jest niezbędny do wzrostu i rozwoju pierwotniaków. Bakteria dostarcza również enzymów cyklu mocznikowego , które są nieobecne u gospodarza. W zamian pierwotniak oferuje swoje enzymy dla pełnych szlaków metabolicznych do biosyntezy aminokwasów , lipidów i nukleotydów , których nie ma w bakterii. Bakteria ma znacznie zredukowany genom w porównaniu z pokrewnymi gatunkami bakterii, brakuje jej wielu genów niezbędnych do jej przeżycia. Fosfatydyloinozytol , lipid błonowy wymagany do interakcji komórka-komórka w bakteriach, jest również syntetyzowany przez pierwotniaka. Bakteria zależy również od gospodarza dla cząsteczek ATP ze względu na swoje funkcje energetyczne. W ten sposób oba organizmy ściśle dzielą i wymieniają swoje systemy metaboliczne.
Kiedy bakteria zostaje zabita przy użyciu antybiotyków , pierwotniak nie może już infekować owadów z powodu zmienionego glikozylofosfatydyloinozytolu (gp63) w wici pierwotniaka. Bezbakteryjny pierwotniak wykazuje zmniejszoną aktywność genów; szczególnie te zaangażowane w proces utleniania-redukcji, transport protonów sprzężony z hydrolizą ATP i glikolizę są zatrzymane. Zmieniają się również elementy strukturalne, w tym powierzchnia komórki, skład węglowodanów, pręcik paraflagellarny i kinetoplast.
Pasożytnictwo
Angomonas deanei została pierwotnie odkryta w przewodzie pokarmowym robaka Zelus leucogrammus . Zdano sobie jednak sprawę, że robaki nie są silnie zainfekowane i prawdopodobnie zostały przeniesione z innych owadów. Obecnie wiadomo, że infekuje różne komary i muchy oraz jest zdolny do infekowania komórek fibroblastów ssaków w warunkach eksperymentalnych. Przenoszenie z jednego owada na drugiego zachodzi tylko między osobnikami dorosłymi ( przenoszenie poziome ), a pierwotniak nie może utrwalić się w jelicie grubym larw owadów. Wić jest używana jako narząd adhezyjny, który przyczepia się w pobliżu gruczołów odbytniczych, a czasem bezpośrednio na powierzchni gruczołów odbytniczych.
Reprodukcja
Rozmnażanie komórkowe wykazuje silną adaptację synergistyczną między bakterią a pierwotniakiem. Najpierw dzieli się bakteria, następnie organelle pierwotniaka, a na końcu jądro . W rezultacie pierwotniaki potomne zawierają dokładnie te same kopie organelli i endosymbiontu bakteryjnego. Cała reprodukcja trwa około 6 godzin w idealnej pożywce hodowlanej; w ten sposób pojedynczy pierwotniak jest w stanie wyprodukować 256 komórek potomnych w ciągu dnia, chociaż może się nieznacznie różnić w swoim naturalnym środowisku.
Endosymbiont i ewolucja
Bakterie symbiotyczne w pierwotniakach trypanosomatycznych pochodzą od β-proteobakterii. W przypadku A. deanei bakterie Ca. „ Kinetoplastibacterium crithidii ” współewoluowały w wzajemnej relacji charakteryzującej się intensywnymi wymianami metabolicznymi. Endosymbiont zawiera enzymy i prekursory metaboliczne, które uzupełniają podstawowe szlaki biosyntezy pierwotniaka gospodarza, takie jak cykl mocznikowy i produkcja heminy i poliaminy .
Bakteria symbiotyczna należy do rodziny β-proteobacterium Alcaligenaceae . Na podstawie genu 16S rRNA wiadomo, że pochodzi on od wspólnego przodka z tym w Strigomonas culicis . Zakłada się, że te dwie grupy wchodzą do dwóch różnych pierwotniaków żywicieli, aby ewoluować w różne gatunki. Stąd nazwa naukowa ( Candidatus ) Kinetoplastybacterium crithidii została nadana bakterii. Chociaż początkowo sugerowano, że bakteria wyewoluowała od wspólnego przodka z przedstawicielami rodzaju Bordetella , szczegółowa analiza filogenomiczna wykazała, że jest ona bliżej spokrewniona z przedstawicielami rodzaju Taylorella . Ponowna analiza przeprowadzona przez GTDB wykazała, że rodzaj jest siostrą Proftelli , symbionta Diaphorina citri .