Poliamina
Poliamina jest związkiem organicznym mającym więcej niż dwie grupy aminowe . Alkilopoliaminy występują naturalnie, ale niektóre są syntetyczne. Alkilopoliaminy są bezbarwne, higroskopijne i rozpuszczalne w wodzie. W pobliżu obojętnego pH występują jako pochodne amonowe. Większość aromatycznych poliamin to krystaliczne ciała stałe w temperaturze pokojowej.
Naturalne poliaminy
Liniowe poliaminy o niskiej masie cząsteczkowej można znaleźć we wszystkich formach życia. Głównymi przykładami są triamina spermidyna i tetraamina spermina . Są one strukturalnie i biosyntetycznie spokrewnione z diaminami, putrescyną i kadaweryną . Metabolizm poliamin jest regulowany przez aktywność enzymu dekarboksylazy ornityny (ODC). Poliaminy występują w dużych stężeniach w mózgu ssaków .
- Naturalne poliaminy
Poliaminy syntetyczne
stanowią ważną z handlowego punktu widzenia klasę syntetycznych poliamin z etylenem ( wiązania -CH2CH2) ; globalne moce produkcyjne oszacowano na 385 000 ton w 2001 roku. Są to półprodukty chemiczne często stosowane do produkcji środków powierzchniowo czynnych oraz jako środki sieciujące żywice epoksydowe . Do interesujących członków tej klasy należą:
- Etylenodiamina , pierwszy członek tej serii. Sam w sobie jest ligandem chelatującym i prekursorem popularnego środka maskującego metale, EDTA (kwasu etylenodiaminotetraoctowego). Permetylowana etylenodiamina daje tetrametyloetylenodiaminę (TMEDA), która ma bardzo wysokie powinowactwo do jonów litu.
- makrocykliczne analogiczne do eterów koronowych : 1,4,7 -triazacyklononan ((NHCH 2CH 2 ) 3 ) i cyklen ((NHCH 2 CH 2 ) 4 ). Pokrewnym makrocyklem tetraaza jest cyklam .
- Tris (2-aminoetylo)amina ( N(CH2CH2NH2 ) 3 ) to rozgałęziona poliamina będąca drugorzędnym produktem ubocznym procesu polietylenoaminowego . Pokrewną trójnożną poliaminą jest 1,1,1-tris (aminometylo) etan . Są to interesujące ligandy chelatujące.
- Polietylenoimina jest polimerem pochodzącym od azyrydyny .
Inne syntetyczne poliaminy obejmują 1,3,5-triazynan (nie mylić z 1,3,5-triazyną ) i N-podstawione analogi. Wiązania metylenowe (-CH2 ) pochodzą od formaldehydu . Produkt reakcji monoetanoloaminy i formaldehydu jest znany w przemyśle jako „triazyna MEA” (w rzeczywistości jest to triazynan) i służy jako rozpuszczalny w wodzie środek usuwający siarkowodór . Heksametylenotetramina (heksamina) to kolejny produkt formaldehydu i amoniaku, który ma różnorodne zastosowania w przemyśle. W kraju wykorzystywany jest jako stałe paliwo kempingowe. W laboratorium reaguje z halogenkami alkilu , selektywnie wytwarzając aminy pierwszorzędowe w reakcji Delépine'a .
- Poliaminy syntetyczne
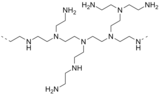
Podjednostka polietylenoiminy
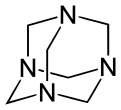
Heksametylenotetramina o strukturze typu adamantanu
amine.svg)


ethane.svg)


Funkcja biologiczna
Chociaż wiadomo, że biosynteza poliamin jest w dużym stopniu regulowana, funkcja biologiczna poliamin jest poznana tylko częściowo. W swojej kationowej formie amoniowej wiążą się z DNA i swoją budową reprezentują związki z kationami występującymi w regularnych odstępach (w przeciwieństwie do Mg 2+
lub Ca 2+
, które są ładunkami punktowymi). Stwierdzono również, że działają one jako promotory programowanego przesunięcia ramki odczytu rybosomów podczas translacji.
Hamowanie biosyntezy poliamin opóźnia lub zatrzymuje wzrost komórek . Dostarczenie egzogennych poliamin przywraca wzrost tych komórek. Większość komórek eukariotycznych wyraża na błonie komórkowej ATPazę przenoszącą poliaminy , która ułatwia internalizację egzogennych poliamin. Układ ten jest bardzo aktywny w szybko proliferujących komórkach i jest celem niektórych obecnie opracowywanych chemioterapeutyków.
Poliaminy są także modulatorami różnych kanałów jonowych , w tym receptorów NMDA i receptorów AMPA . Blokują skierowane do wewnątrz kanały potasowe prostownika, tak że prądy kanałów są prostowane do wewnątrz, w ten sposób zachowana jest energia komórkowa, tj. gradient jonów K +
przez błonę komórkową. Ponadto poliamina uczestniczy w inicjowaniu ekspresji odpowiedzi SOS operonu Colicyny E7 i reguluje w dół białka niezbędne do wychwytu kolicyny E7, zapewniając w ten sposób przewagę w przeżywalności E. coli wytwarzającym kolicynę w warunkach stresowych.
Poliaminy mogą zwiększać przepuszczalność bariery krew-mózg .
Biorą udział w modulowaniu starzenia się narządów roślin i dlatego są uważane za hormon roślinny . Ponadto biorą bezpośredni udział w regulacji programowanej śmierci komórki.
Naprawa DNA ukierunkowana na homologię
Poliaminy wspomagają naprawę pęknięć dwuniciowych (DSB) za pośrednictwem rekombinacji homologicznej (HR) . Poliaminy zwiększają aktywność wymiany nici DNA rekombinazy RAD51 . Wyczerpywanie się poliamin uwrażliwia komórki na substancje genotoksyczne, takie jak promieniowanie jonizujące i ultrafiolet promieniowanie. Wpływ poliamin na RAD51 wynika z ich zdolności do wzmacniania wychwytywania homologicznego dupleksu DNA i promowania parowania i wymiany homologicznego DNA za pośrednictwem RAD-51. Wydaje się, że poliaminy odgrywają utrwaloną ewolucyjnie rolę w regulowaniu aktywności rekombinazy.
Biosynteza spermidyny, sperminy, termosperminy

Spermidyna jest syntetyzowana z putrescyny przy użyciu grupy aminopropylowej z dekarboksylowanej S -adenozylo-L-metioniny (SAM). Reakcja jest katalizowana przez syntazę spermidyny .
Sperma syntetyzowana jest w reakcji spermidyny z SAM w obecności enzymu syntazy sperminy .
Poliaminy ulegają szybkiej przemianie wzajemnej w cyklu poliaminowym, w którym putrescyna prowadzi do syntezy spermidyny i sperminy, z degradacją tych poliamin do putrescyny, która może rozpocząć cykl od nowa.
Termospermina (NH 2 -(CH 2 ) 3 -NH-(CH 2 ) 3 -NH-(CH 2 ) 4 -NH 2 ) jest strukturalnym izomerem sperminy i nowym typem regulatora wzrostu roślin. Jest wytwarzany ze spermidyny w wyniku działania syntazy termosperminy , która jest kodowana przez gen o nazwie ACAULIS5 (ACL5).
Analogi poliamin
Krytyczna rola poliamin we wzroście komórek doprowadziła do opracowania szeregu środków zakłócających metabolizm poliamin. Środki te są stosowane w terapii nowotworów. Analogi poliamin zwiększają poziom p53 w komórce, co prowadzi do ograniczenia proliferacji i apoptozy. Zmniejsza także ekspresję receptora estrogenowego alfa w raku piersi ER-dodatnim.
Linki zewnętrzne
- Poliaminy w proliferacji cyklu komórkowego i śmierci komórek
- Dekarboksylaza ornityny: ekspresja i regulacja w mózgu szczura i u myszy transgenicznych, 2002, Pekka Kilpelainen, Wydział Biochemii, Uniwersytet w Oulu. Obszerny przegląd literatury do roku 2001 na temat struktury, właściwości, metabolizmu poliamin u ssaków oraz roli fizjologicznej i patofizjologicznej poliamin (zobacz spis treści artykułu)