N-acetylotransferaza fosforanowa glukozaminy
| N-acetylotransferaza 6-fosforanu | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 glukozaminy N-acetylotransferaza 6-fosforanu glukozaminy 1, homodimer, ludzkie
| |||||||||
| identyfikatory | |||||||||
| nr WE | 2.3.1.4 | ||||||||
| nr CAS | 9031-91-8 | ||||||||
| Bazy danych | |||||||||
| IntEnz | Widok IntEnz | ||||||||
| BRENDA | Wpis BRENDY | ||||||||
| ExPASy | Widok NiceZyme | ||||||||
| KEGG | Wpis KEGG | ||||||||
| MetaCyc | szlak metaboliczny | ||||||||
| PRYM | profil | ||||||||
| Struktury PDB | RCSB PDB PDBe PDB suma | ||||||||
| Ontologia genów | AmiGO / QuickGO | ||||||||
| |||||||||
W enzymologii N-acetylotransferaza glukozaminofosforanowa (GNA) ( EC 2.3.1.4 ) jest enzymem , który katalizuje przeniesienie grupy acetylowej z acetylo-CoA do pierwszorzędowej aminy w glukozamido-6-fosforanie, generując wolny CoA i N -acetylo-D-glukozamino-6-fosforan.
Enzym ten należy do rodziny transferaz , grupy enzymów, które przenoszą bardzo specyficzną grupę funkcyjną, w tym przypadku acetyl, od dawcy do receptora. W szczególności enzym ten można scharakteryzować jako część rodziny acylotransferaz , ponieważ obejmuje on przeniesienie ogólnej grupy acylowej z metylem jako podstawnikiem.
Nomenklatura
Systematyczna nazwa tej klasy enzymów to N-acetylotransferaza acetylo-CoA:D-glukozamino-6-fosforanowa. Inne powszechnie używane nazwy to transacetylaza fosfoglukozaminy, acetylaza fosfoglukozaminy, acetylaza glukozamino-6-fosforanowa, D-glukozamino-6-P N-acetylotransferaza, acetylotransferaza aminodeoksyglukozofosforanowa, acetylaza glukozamino-6-fosforanowa, N-acetylotransferaza glukozamino-6-fosforanowa, N-acetyloglukozamina Syntaza -6-fosforanowa, N-acetylaza fosfoglukozaminy, N-acetylotransferaza glukozaminofosforanowa i N-acetylotransferaza glukozamino-6-fosforanowa.
Funkcjonować
Enzym ten jest częścią szlaku biosyntezy heksozaminy (HBP), który jest jednym ze szlaków przetwarzania glukozy w ogólnym metabolizmie. Szlak ten ma wspólne dwa początkowe etapy z glikolizą i różni się tylko niewielką częścią strumienia glukozy od tego bardziej tradycyjnego szlaku glikolitycznego. Dlatego korzystne jest, gdy istnieje regulacja ujemnego sprzężenia zwrotnego na glikolizę, jak w przypadku dużych ilości wolnych kwasów tłuszczowych . Produktem końcowym tego szlaku jest UDP-N-acetyloglukozamina , która bierze udział w modyfikacji złożonych cząsteczek, takich jak glikolipidy , proteoglikany i glikoproteiny . Ten produkt końcowy działa jako nośnik N-acetyloglukozaminy , która jest monomeryczną jednostką chityny, polimeru strukturalnego, z którego składają się skorupy skorupiaków i owadów , a także ściana komórkowa grzybów . Ponadto N-acetyloglukozamina jest również jednostką polimeru peptydoglikanu , który tworzy ścianę komórkową bakterii wraz z disacharydem N-acetylomuraminowym.

Mówiąc dokładniej, enzym GNA katalizuje czwarty etap szlaku HBP u eukariontów , promując transfer węgla z acetylo-CoA na inny substrat, D-glukozamino-6-fosforan, który ostatecznie daje UDP-N-acetyloglukozaminę . Jest to mały, ale ważny krok chemiczny, który ma kluczowe znaczenie dla właściwości subproduktów tego szlaku metabolicznego. Acetylowanie prowadzi się aż do samego produktu końcowego szlaku heksaminy i jest bardzo charakterystyczne dla polimerów utworzonych z N -acetyloglukozaminy . Na przykład stanowi jedną z głównych różnic w strukturze molekularnej chityny i celulozy i wyjaśnia wiele fizycznych i chemicznych właściwości tych polimerów. Na przykład w przypadku chityny badania obliczeniowe wykazały, że acylowanie przyczynia się do tworzenia wiązań wodorowych , które stabilizują strukturę krystaliczną tego polimeru, zapewniając większą odporność na pękanie.
Niemniej jednak w metabolizmie prokariotycznym szlak biosyntezy heksozaminy przebiega przez inny etap reakcji, w którym inny enzym działa na te same charakterystyczne substraty (ryc. 1). U prokariotów przeniesienie fosforanu z węgla 6 na węgiel 1 odbywa się przed acylowaniem, tak że substratem reakcji dodawania węgla jest raczej glukozamino-1-fosforan niż 6-fosforan D-glukozaminy. Tym razem enzymem odpowiedzialnym za acetylację jest dwufunkcyjne białko glmU (N-acetyloglukozamino-1-fosforano-urydylotransferaza), które katalizuje również dodanie UDP do grupy fosforanowej na N-acetylo-D-glukozamino-1-fosforanie.
U ludzi N-acetylotransferaza glukozaminofosforanowa jest dimerem z dwiema identycznymi podjednostkami i jest kodowana w genie GNPNAT (symbol HGNC). Dokładniej, enzym ulega silnej ekspresji w wątrobie , żołądku i tkankach przewodu pokarmowego, a wewnątrz komórki jest zlokalizowany w endosomach iw aparacie Golgiego (według ręcznej adnotacji).
Mechanizm
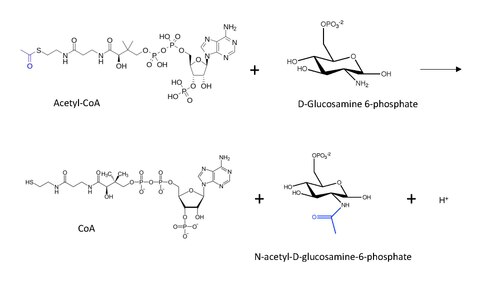
Strukturę molekularną reakcji katalizowanej przez GNA pokazano poniżej, z przeniesioną grupą acetylową zaznaczoną na niebiesko.
Ogólny mechanizm reakcji postulowany dla acetylacji N-końca białka (inspirowany mechanizmem acetylacji lizyny) z Acetylo-CoA obejmuje atak nukleofilowy grupy aminowej (w tym przypadku z D-glukozamino-6-fosforanu) na końcowy karbonyl w węglu transfer, co prowadzi do powstania czworościennego związku pośredniego węgla. Reakcja przebiega z przywróceniem karbonylu przez usunięcie CoA jako grupy opuszczającej, tak że teraz grupa acetylowa jest połączona z grupą aminową w drugim substracie.
Specjalnie dla tych katalizatorów N-acetylotransferazy, badania z enzymem GNA S. cerevisiae wykazały, że niektóre specyficzne aminokwasy przyczyniają się do wiązania substratu, zwiększonej nukleofilowości grupy aminowej i ostatecznie katalizy, co wspiera postulowany mechanizm opisany powyżej. Glu98, Asp99 i Ile100 polaryzują karbonylowe w Acetylo-CoA , zwiększając elektrofilowość węgla , jak również stabilizując tetraedryczny związek pośredni węgla. Tyr143 odpowiada za stabilizację tiolanu anion, faworyzując S-CoA jako grupę opuszczającą z tetraedrycznego węgla. Wreszcie, Asp134 zwiększa nukleofilowość grupy aminowej w D-glukozamido-6-fosforanie poprzez przekazanie gęstości elektronowej atomowi azotu . W innym organizmie, C. albicans , stwierdzono, że podobny zestaw aminokwasów jest niezbędny do aktywności katalitycznej, odpowiednio układ Glu88-Asp-89-Ile90, Asp125 i Tyr133.
Struktura

Pod koniec 2019 roku rozwiązano 13 struktur dla tej klasy enzymów u różnych gatunków, z kodami dostępu PDB 1I12 ( Saccharomyces cerevisiae ), 1I1D ( Saccharomyces cerevisiae ), 1I21 ( Saccharomyces cerevisiae ), 2HUZ ( Homo sapiens ), 2O28 ( Homo sapiens ), 4AG7 ( Caenorhabditis elegans ), między innymi.
Rycina 3 przedstawia proponowaną strukturę krystaliczną GNA u ludzi , z każdą podjednostką katalityczną w innym kolorze. Acetylo -CoA związany z enzymem jest pokazany na jasnoróżowo, a produkt nadal związany z miejscem katalitycznym jest pokazany na fioletowo. Przeniesiona acetylowa w produkcie N-acetylo-D-glukozamino-6-fosforanowym na fioletowo jest pokazana na żółto. Ta proponowana struktura 3D białka pokazuje , że określone części substratów biorących udział w tej reakcji - końcowy koniec liniowej części Acetylo-CoA i azot grupy przyłączone do pierścienia glukozaminy - są bardzo blisko.