Lista makrocząsteczkowych struktur krystalicznych o znaczeniu biofizycznym

Struktury krystaliczne cząsteczek białek i kwasów nukleinowych oraz ich kompleksów mają kluczowe znaczenie dla praktyki większości działów biofizyki i ukształtowały wiele z tego, co rozumiemy naukowo na poziomie szczegółowości atomowej biologii. Ich znaczenie podkreśla Organizacja Narodów Zjednoczonych, ogłaszając rok 2014 Międzynarodowym Rokiem Krystalografii , jako setną rocznicę przyznania przez Maxa von Laue w 1914 roku nagrody Nobla za odkrycie dyfrakcji promieni rentgenowskich przez kryształy. Ta chronologiczna lista ważnych biofizycznie struktur białek i kwasów nukleinowych jest luźno oparta na przeglądzie w Biophysical Journal . Lista obejmuje wszystkie pierwsze tuziny odrębnych struktur, te, które otworzyły nowe możliwości w zakresie przedmiotu lub metody, oraz te, które stały się systemami modelowymi do pracy w przyszłych biofizycznych obszarach badań.
mioglobina


1960 - Mioglobina była pierwszą strukturą krystaliczną cząsteczki białka o wysokiej rozdzielczości. Mioglobina kołysze zawierającą żelazo hemową , która odwracalnie wiąże tlen do zasilania włókien mięśniowych , a te pierwsze kryształy pochodziły z mioglobiny z kaszalota , którego mięśnie potrzebują obfitego magazynowania tlenu do głębokich nurkowań. Trójwymiarowa struktura mioglobiny składa się z 8 helis alfa , a struktura krystaliczna wykazała, że ich konformacja była prawoskrętna i bardzo ściśle odpowiadała geometrii zaproponowanej przez Linusa Paulinga , z 3,6 resztami na obrót i szkieletowymi wiązaniami wodorowymi od peptydu NH jednej reszty do peptydu CO reszty i+4. Mioglobina jest układem modelowym dla wielu rodzajów badań biofizycznych, zwłaszcza obejmujących proces wiązania małych ligandów, takich jak tlen i tlenek węgla .
Hemoglobina
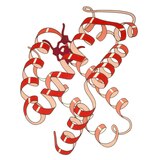

1960 - Struktura krystaliczna hemoglobiny wykazała tetramer dwóch pokrewnych typów łańcuchów i została rozwiązana w znacznie niższej rozdzielczości niż monomeryczna mioglobina, ale wyraźnie miała tę samą podstawową architekturę 8-helisową (obecnie nazywaną „fałdem globiny”). Dalsze struktury krystaliczne hemoglobiny w wyższej rozdzielczości [PDB 1MHB, 1DHB) wkrótce wykazały sprzężoną zmianę zarówno lokalnej, jak i czwartorzędowej konformacji między stanami oksy i dezoksy hemoglobiny, co wyjaśnia współdziałanie wiązania tlenu we krwi i allosteryczne działanie czynników takich jak jak pH i DPG . Przez dziesięciolecia hemoglobina była głównym przykładem nauczania koncepcji allostery, a także była przedmiotem intensywnych badań i dyskusji na temat allostery. W 1909 roku kryształy hemoglobiny z ponad 100 gatunków zostały użyte do powiązania taksonomii z właściwościami molekularnymi. Ta książka została zacytowana przez Perutza w raporcie z 1938 roku o kryształach hemoglobiny końskiej, który rozpoczął jego długą sagę mającą na celu rozwiązanie struktury kryształu. Kryształy hemoglobiny są pleochroiczne - ciemnoczerwone w dwóch kierunkach i bladoczerwone w trzecim - z powodu orientacji hemów i jasnego pasma Soreta porfiryny hemu grup jest używany w analizie spektroskopowej wiązania ligandu hemoglobiny.

Lizozym z białka jaja kurzego
1965 - Lizozym białka jaja kurzego (plik PDB 1lyz). była pierwszą strukturą krystaliczną enzymu (rozszczepia małe węglowodany na cukry proste), stosowaną do wczesnych badań mechanizmu działania enzymu. Zawierał arkusz beta (antyrównoległy) oraz helisy, a także był pierwszą strukturą makrocząsteczkową, której współrzędne atomowe zostały udoskonalone (w rzeczywistej przestrzeni). Surowiec do przygotowania można kupić w sklepie spożywczym, a lizozym z jaja kurzego bardzo łatwo krystalizuje w wielu różnych grupach przestrzennych ; jest to ulubiony przypadek testowy dla nowych eksperymentów i instrumentów krystalograficznych. Niedawnymi przykładami są nanokryształy lizozymu do gromadzenia danych laserowych na swobodnych elektronach oraz mikrokryształy do dyfrakcji mikroelektronów.

Rybonukleaza
1967 - Rybonukleaza A (plik PDB 2RSA) jest enzymem rozszczepiającym RNA, stabilizowanym 4 wiązaniami dwusiarczkowymi. Został użyty w Anfinsena nad fałdowaniem białek, które doprowadziły do koncepcji, że trójwymiarowa struktura białka jest określona przez jego sekwencję aminokwasową . Rybonukleaza S , rozszczepiona, dwuskładnikowa forma badana przez Freda Richardsa , był również aktywny enzymatycznie, miał prawie identyczną strukturę krystaliczną (plik PDB 1RNS) i wykazano, że jest aktywny katalitycznie nawet w krysztale, pomagając rozwiać wątpliwości co do znaczenia struktur krystalicznych białek dla funkcji biologicznych.

Proteazy serynowe
1967 - Proteazy serynowe są historycznie bardzo ważną grupą struktur enzymatycznych, ponieważ wspólnie oświetliły mechanizm katalityczny (w ich przypadku przez „triadę katalityczną Ser-His-Asp”), podstawę różnych specyficzności substratów i mechanizm aktywacji dzięki któremu kontrolowane rozszczepienie enzymatyczne zakopuje nowy koniec łańcucha, aby odpowiednio zmienić miejsce aktywne. Wczesne struktury krystaliczne obejmowały chymotrypsynę (plik PDB 2CHA), chymotrypsynogen (plik PDB 1CHG), trypsynę (plik PDB 1PTN) i elastazę (Plik PDB 1EST). Były też pierwszymi strukturami białkowymi, które wykazywały dwie niemal identyczne domeny, przypuszczalnie spokrewnione przez duplikację genów . Jednym z powodów ich szerokiego zastosowania jako przykładów w podręcznikach i klasach był system numeracji kodu wstawiania (znienawidzony przez wszystkich programistów komputerowych), który sprawił, że Ser195 i His57 były spójne i zapadające w pamięć pomimo różnic w sekwencjach specyficznych dla białek.
Papaina
1968 - Papaina


Karboksypeptydaza
1969 - Karboksypeptydaza A jest metaloproteazą cynku . Jego struktura krystaliczna (plik PDB 1CPA) wykazywała pierwszą równoległą strukturę beta: duży, skręcony, centralny arkusz 8 nici z aktywnym miejscem Zn zlokalizowanym na C-końcu środkowych nici i arkusz otoczony z obu stron przez helisy alfa. Jest to egzopeptydaza który rozszczepia peptydy lub białka od końca karboksylowego, a nie od wewnątrz sekwencji. Później opracowano mały białkowy inhibitor karboksypeptydazy (plik PDB 4CPA), który mechanicznie zatrzymuje katalizę, prezentując swój C-końcowy koniec wystający spomiędzy pierścienia wiązań dwusiarczkowych ze zwartą strukturą za nim, uniemożliwiając enzymowi zassanie łańcucha poza pierwszą pozostałość.

Subtylizyna
1969 - Subtylizyna (plik PDB 1sbt) była drugim typem proteazy serynowej z niemal identycznym miejscem aktywnym jak enzymy z rodziny trypsyny, ale z zupełnie innym ogólnym fałdem. Dało to pierwszy pogląd na zbieżną ewolucję na poziomie atomowym. Później intensywne badanie mutacji subtylizyny udokumentowało wpływ wszystkich 19 innych aminokwasów na każdą pojedynczą pozycję.
Dehydrogenaza mleczanowa
1970 - Dehydrogenaza mleczanowa

Inhibitor trypsyny
1970 - Podstawowy trzustkowy inhibitor trypsyny lub BPTI (plik PDB 2pti) jest małym, bardzo stabilnym białkiem, które było wysoce produktywnym systemem modelowym do badań supermocnego wiązania, tworzenia wiązań dwusiarczkowych (SS), fałdowania białek , stabilności molekularnej przez mutacje aminokwasów lub wymianę wodór-deuter i szybką dynamikę lokalną przez NMR . Biologicznie BPTI wiąże i hamuje trypsynę podczas przechowywania w trzustce , umożliwiając aktywację trawienia białka dopiero po uwolnieniu trypsyny do żołądka.
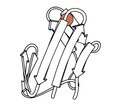
Rubredoksyna
1970 - Rubredoksyna (plik PDB 2rxn) była pierwszą rozwiązaną strukturą redoks, minimalistycznym białkiem z żelazem związanym 4 łańcuchami bocznymi Cys z 2 pętli na szczycie spinek do włosów β. Ulegał dyfrakcji do 1,2Å, umożliwiając pierwsze udoskonalenie białka w przestrzeni odwrotnej (4,5rxn). [Uwaga: uważaj 4rxn, zrobione bez ograniczeń geometrii!] archeologiczne stanowią wiele małych struktur o najwyższej rozdzielczości w PDB.


Insulina
1971 - Insulina (plik PDB 1INS) jest hormonem odpowiedzialnym za metabolizm cukru i magazynowanie tłuszczu oraz ma znaczenie w chorobach człowieka, takich jak otyłość i cukrzyca . Jest biofizycznie godny uwagi ze względu na wiązanie Zn, równowagę między stanami monomeru, dimeru i heksameru, zdolność do tworzenia kryształów in vivo i syntezę jako dłuższej formy „pro”, która jest następnie rozszczepiana, aby złożyć się jako aktywna 2- łańcuch, monomer połączony z SS. Insulina była sukcesem programu wzrostu kryształów NASA na promie kosmicznym , wytwarzając preparaty masowe o bardzo jednolitych drobnych kryształach do kontrolowanego dawkowania.
Nukleaza gronkowcowa
1971 - Nukleaza gronkowcowa
Cytochrom C
1971 - Cytochrom C
Lizozym faga T4
1974 - Lizozym faga T4
Immunoglobuliny
1974 - Immunoglobuliny
Dysmutaza ponadtlenkowa
1975 - Dysmutaza ponadtlenkowa Cu,Zn
Przenieś RNA
1976 - Transfer RNA

Izomeraza triozofosforanowa
1976 - Izomeraza fosforanu triozy
Pepsynopodobne proteazy asparaginianowe
1976 - Rhizopuspepsyna
1976 - Endotiapepsyna
1976 - penicylopepsyna
Późniejsze struktury (od 1978 r.)
1978 - Wirus dwudziestościenny
1981 - Dodekamer DNA w formie B Dickersona
1981 - Rak
1985 - Kalmodulina
1985 - polimeraza DNA

1985 — Centrum reakcji fotosyntezy Pary bakteriochlorofilów (zielone) wewnątrz błony wychwytują energię ze światła słonecznego, a następnie pokonują wiele etapów, by stać się dostępnymi w grupach hemowych (czerwone) w module cytochromu-C na górze. Była to pierwsza struktura krystaliczna rozwiązana dla białka błonowego, kamień milowy uznany przez Nagrodę Nobla dla Hartmuta Michela, Hansa Deisenhofera i Roberta Hubera.
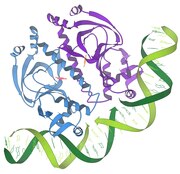
1986 - Interakcje represor/DNA
1987 - Główny kompleks zgodności tkankowej "
1987 - Ubikwityna
1987 - Białko ROP
1989 - proteaza HIV-1
1990 - Bakteriorodopsyna
1991 - Cewka spiralna GCN4
1991 - odwrotna transkryptaza HIV-1
1993 - Beta helisa liazy pektynianowej
1994 - Kolagen
1994 - Kompleks Barnase / Barstar
1994 - ATPaza F1
1995 - Heterotrimeryczne białka G
1996 - Zielone białko fluorescencyjne

1996 - Białko motoryczne kinezyny
1997 - GroEL /ES opiekun
1997 - Nukleosom
1998 - Intron samoskładający się grupy I

1998 - Topoizomerazy DNA wykonują biologicznie ważną i niezbędną pracę polegającą na rozplątywaniu nici DNA lub helis, które splatają się ze sobą lub są zbyt mocno skręcone podczas normalnych procesów komórkowych, takich jak transkrypcja informacji genetycznej .
1998 - Tubulina dimer alfa/beta
1998 - Kanał potasowy
1998 - skrzyżowanie Holliday
2000 - Rybosomy są centralną częścią biologii i biofizyki, która po raz pierwszy stała się dostępna strukturalnie w 2000 roku.

2000 - ATPaza AAA+
2002 - Ankyrin powtarza
2003 - projekt białka TOP7
2004 - Cyjanobakteryjne białka zegara dobowego
2004 - Przełącznik rybny
2006 - Ludzki egzosom

2007 - Receptor sprzężony z białkiem G

2009 - Cząsteczka Vault jest intrygującym nowym odkryciem dużej pustej w środku cząstki, powszechnej w komórkach, z kilkoma różnymi sugestiami dotyczącymi jej możliwej funkcji biologicznej. Struktury krystaliczne (pliki PDB 2zuo, 2zv4, 2zv5 i 4hl8) pokazują, że każda połowa sklepienia składa się z 39 kopii długiego 12-domenowego białka, które wirują razem, tworząc obudowę. Nieporządek na samej górze i dole sugeruje otwory umożliwiające dostęp do wnętrza sklepienia.
